In general, eggs are produced every other moulting period (Joosse & Veltkamp, 1970).
Egg production shows a clear relationship with moulting.
Since the moulting process is temperature dependent it thus can be expected
that egg laying will be more frequent at higher temperatures, due to the
increased moulting rate (Joosse & Veltkamp, 1970).
In Isotoma viridis, the number of egg laying periods being reported
as 3 by Hale (1965), and a maximum of 8 by Joosse & Veltkamp (1970).
In Isotoma viridis, a mature female lays one batch of 27-54 eggs
(after Milne, 1960).
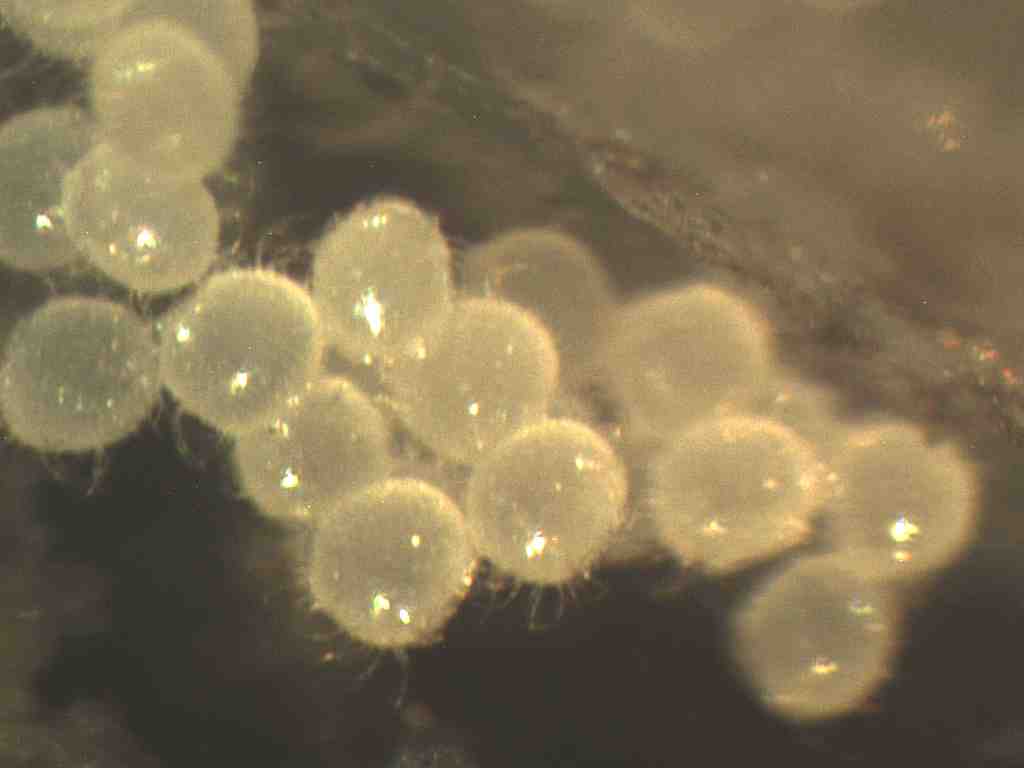
The eggs of Ceratophysella armata are laid in small clumps in damp
situations under surface of loose stones, wood, or leaves (Tiegs, 1942:154).
Sminthurus viridis attains sexual maturity during the sixth instar when
oviposition may already take place. On very wet soil the eggs are laid on
elevations and surface litter. On dry soil the eggs are deposited in more
moist cracks. The female eats soil that is being excreted as a sticky mixture
which is used to cover the eggs.
The eggs are laid in batches.
The female stands with widespread legs and elevated abdomen (fig. 000).
A drop of soil excreta is slowly forced out of the anus, followed by the
extrusion from the gonopore of the shapeless egg which quickly assumes a
spherical form.
The abdomen is then depressed and the egg is firmly pressed against the soil
by means of a cicular movement of the abdomen, the excreta serving to anchor
the egg.
A further drop of soil excreta is then spread over the egg.
Uncovered eggs may be eaten by the female or other springtails.
Oviposition may take several minutes for each egg.
12 hours may be spent on laying a single egg batch.
Oviposition may commence during the sixth instar and continue up to the
tenth instar.
A female lays about 15 to 87 eggs during her lifetime.
Egg batches of several different females may be clustered (up to 700 eggs
in laboratory conditions).
(After Walters, 1967:23-24).
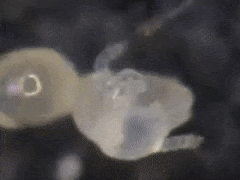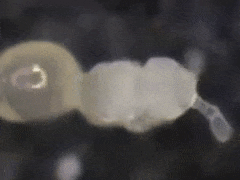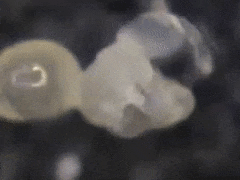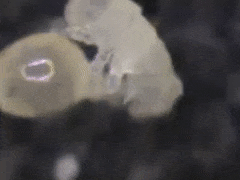
In Sminthurides aquaticus eggs are deposited in cuplike depressions
of the epidermis of Lemna minor (Fig.001).
The cups have been
chewn out
by the female.
After deposition, the egg is
covered by a faecal pellet.
In Sinella curviseta, a typical female of moderate age
(i.e., more than 4 weeks after eclosion)
deposits clutches averaging 59 eggs each,
at about 8-day intervals (Waldorf 1971:673).
About 10 hours are required for the deposition of a clutch (Waldorf 1971:674).
Approximately 2% of each clutch fails to develop;
these eggs remain smooth after chorion rupture while in developing eggs
of the same clutch the rupture gives them the rough surface (Waldorf 1971:674).
About 24 hours are necessary for an egg to develop a rough surface (Waldorf 1971:674).
"La forme de l'oeuf, sa couleur et le tissu de l'enveloppe varient non
seulement d'un genre à l'autre, mais encore d'espèce à
espèce; les oeufs à enveloppe solide sont en
général très peu transparents, lisses, d'une couleur
brune plus ou moins foncée et plus souvent oblongs que
sphériques: ils appartiennent surtout au genre Podure.
Ceux à enveloppe molle offrent plus de variété dans la
forme et la texture de la membrane extérieure;
ils sont tantôt oblongs ou ovoïdes, tantôt sphériques
ou en sphéroïde aplati de deux côtés; leur couleur
est généralement pâle, ou plutôt blanche, mais
légèrement lavée de blue, de jaune, de rose ou de violet,
selon les espèces. Leur transparence permet de suivre jusqu'à
un certain point le développement de l'embryon. La membrane
extérieure est lisse dans la plupart des espèces,
pointillée ou réticulée dans quelques unes; dans ce
dernier cas, les oeufs sont parfois velus; garnis de poils longs et
serrés; d'autres sont plutôt épineux que velus; les
épines longues, flexibles et un peu frisées comme de la laine,
affectant toutes les formes et toutes les directions, sont larges à
leur base et aiguës à leur extrémité; elles
naissent chacune d'une espèce de bulbe formé de deux
renflements placés l'un au dessus de l'autre, dont le premier ou
l'inférieur est hémisphérique, et le second ou le
supérieur, en disque arrondi; c'est au centre de ce dernier qu'est
attachée l'épine.
Dans les oeufs réticulés et velus, c'est du point de jonction
des lignes qui forment les mailles du réseau, que sortent les poils;
ils sont droits ou perpendiculaires au centre de l'oeuf et n'ont jamais de
bulbe pour base."
(Nicolet, 1842:16-17).
Egg sizes of many species are between 0.12 and 0.20 mm (Anderson, 1973:178).
Some of the larger are
Sminthurinus mime : 0.25 mm (Ashraf, 1969 cited from Anderson, 1973:178),
Sminthurus viridis : 0.27 mm (Tiegs, 1942 cited from Anderson, 1973:178),
Entomobrya marginata : 0.27 mm (Tiegs, 1942 cited from Anderson, 1973:178),
Anurida maritima : 0.27 mm (Claypole, 1898 cited from Anderson, 1973:178),
Pogonognathellus longicornis : 0.37 mm (Uzel, 1898 cited from Anderson, 1973:178 as Tomocerus plumbeus),
Blasconura hirtella : 0.42 mm (Tiegs, 1942 cited from Anderson, 1973:178 as Achorutes hirtellus),
Tetrodontophora bielanensis : 0.50 mm (Jura, 1965 cited from Anderson, 1973:178).
The eggs of Ceratophysella armata are spherical measuring not more
than 0.12 mm in diameter (Tiegs, 1942:154).
The eggs of Isotoma viridis are smooth surfaced and globular and
measure 0.21 mm in diameter before development (Milne, 1960).
Oviposition always occurs on the surface and on laying the eggs have an
orange-red pigmentation (Milne, 1960).
The eggs of Sminthurus viridis are spherical and measure about 0.27 mm
in diameter (Tiegs, 1942:162).
The eggs of Sinella curviseta from deposition to about 24 hours of age
are smooth shining, and spherical;
from 24 hours of age until hatching (about 5 days after deposition),
they are rough, dull, and slightly elongate.
Smooth eggs include very young eggs
and faulty eggs of any age (Waldorf, 1971:673).
At oviposition (fig.1, 1b), many eggs of Collembola
are spherical and smooth (Lubbock, 1873:82).
The eggs have a smooth external shell (Kühnelt, 1961:162),
called the 'chorion' (Wigglesworth, 1965:1).
The egg is enclosed in an external chorion secreted by the follicle cells
of the parent ovariole
(Anderson, 1973:176).
E.g., the eggs of Isotoma Walkerii(sic) (now Desoria albella)
are spherical, glistening white, with the chorion very transparent.
(Packard, 1871? cited from Lubbock 1873:84).
In Hypogastruridae, the eggs are spherical, whitish, opaque and
shiny; the chorion is smooth, in general; the egg is moist and sticky
(Thibaud, 1970:127).
In Folsomia candida, the fresh egg is milky white and has a diameter
of 0.110-0.126 mm; the surface of the egg shell is finely granulated
(Gao, Bu, Luan & Yin, 2006:519).
In Hypogastruridae, at oviposition, the egg diameter is about 140-260 micron
(Thibaud, 1970:128).
In subterraneous species, the number of eggs per batch decreases, while the
size of the eggs increases
(Thibaud, 1970:130).
In Hypogastruridae, the incubation period (time between oviposition and
eclosion) is about 24-45 days at 9-12°C
(Thibaud, 1970:130).
In Folsomia candida, the incubation period is about 7 to 10 days
(Gao, Bu, Luan & Yin, 2006:519).
In Megalothorax, the incubation period is about 16 days at 20°C
(Blancquaert & Mertens, 1979:126).
In Hypogastruridae, the eggs can develop and hatch under water
(Thibaud, 1970:181).
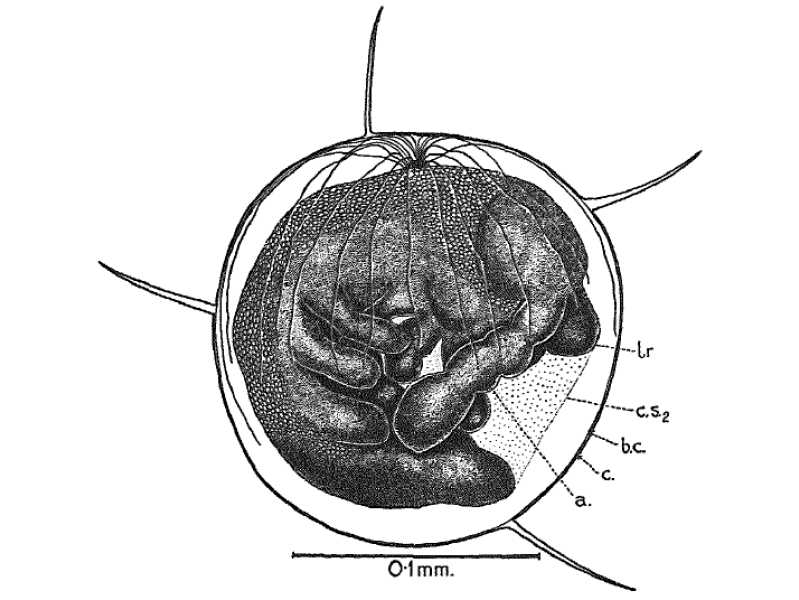
Fig.1c. Egg of Pseudosinella alba After Tiegs, 1942:161 Text-Fig.3
The chorion of the egg in
Lepidocyrtus (Fig.1cc) and in
Pseudosinella alba is furnished with long
delicate spines in the early stage of ventral flexure of the embryo
when the chorion is not yet ruptured (Fig.1c) (Tiegs, 1942:160-161).
The spherical egg has a central nucleus in a small cytoplasmic halo,
surrounded by uniformly distributed yolk granules in a
sparse cytoplasmic reticulum
(after Anderson, 1973:176,178).
Due to the formation of a dorsal organ,
the form of the egg changes by the rising of the dorsum of the growing embryo
(Schaller, 1970:51).
The dorsal organ consists of filamentous processes arising from a pit in the
anterior dorsal region that are grown on to the lower pole of the egg
(Tiegs, 1942:161) (Wigglesworth, 1965:5).
It is suggested that these filaments may be concerned in the absorption of water
(Tiegs, 1942:165-166) (Wigglesworth, 1965:5).
The egg swells soon after deposition
(Davidson, 1932:867; Schaller, 1970:51; Thibaud, 1970:130-132),
due to absorption of water (Thibaud, 1970:130).
The eggs do not remain spherical at all; they increase in size rapidly after
oviposition, to remain stable halfway the incubation period
(Thibaud, 1970:130-131).
As a result, usually on the second to fourth day of embryonic development,
the chorion bursts (Schaller, 1970:51).
And the blastodermic cuticle takes it place as the protecting sheath for
the embryo (Tiegs, 1942:156).
The egg of Ceratophysella armata, now enlarged, measures 0.17 mm
in diameter (Tiegs, 1942:156).
The rupture of the chorion in Sminthurus viridis takes place
later than in Ceratophysella (Tiegs, 1942:162).
In many species, the chorion does not split randomly,
but symmetrically at the equator of the egg
(Davidson, 1932:867; Schaller, 1970:51; Thibaud, 1970:132).
The ruptured chorion adheres in two pieces to the blastodermic cuticle
(Tiegs, 1942:156).
The two chorion caps thus formed,
due to the swelling of the egg,
gradually separate to form two symmetric caps at the poles of the egg,
the 'polar caps' (fig.2)
(Nicolet, 1842:19; Davidson, 1932:867; Schaller, 1970:51).
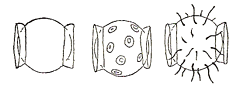
Fig.2. Polar caps in eggs of Collembola:
Protaphorura armata (left)
Deuteraphorura inermis (middle)
Orchesella cincta (right)
After Handschin in Kühnelt, 1961:162
Nicolet, 1842:19 is the first to describe the forming of polar caps (fig.6):
"Dès que les yeux commençent à paraître, l'oeuf se
comprime de manière à prendre la forme d'un sphéroïde
aplati; l'enveloppe extérieure se fend dans son plus grand
diamètre et forme deux hémisphères qui s'éloignent
insensiblement l'un de l'autre; leur sommet s'affaisse, et leur donne
bientôt l'apparence de bonnets grecs, appliqués de chaque
côté de la nouvelle membrane, qui devient alors, jusqu'à
l'éclosion, membrane extérieure de l'oeuf."
Directly as the eggs of Isotoma viridis are laid they begin to enlarge
due to uptake of water from the atmosphere. In an atmosphere of 100%
relative humidity eggs reach maximum size in a few hours. Further
enlargement does not take place until the growth of the embryo begins
after about 20% of the developmental period (Hale, 1965c).
The eggs become coloured due to the formation of pigment in the body of
the embryo, however eye spots appear before the body colouration.
Development can be characterized as epimetabolic (Weber, 1968),
which means a gradual differentiation occurs during development.
The separating chorion caps reveal the
embryo surrounded by a transparant membraneous inner shell
(called the 'vitelline membrane' (Wigglesworth, 1965:1))
(Davidson, 1932:867; Thibaud, 1970:132).
In Hypogastruridae, between the 12th to 15th day, the limbs can be recognised
through the transparant vitelline membrane;
around the 16th day, in pigmented species, the two ocular patches can be seen
(Thibaud, 1970:132) (fig.3a, 3aa, 3ab, 3ac).
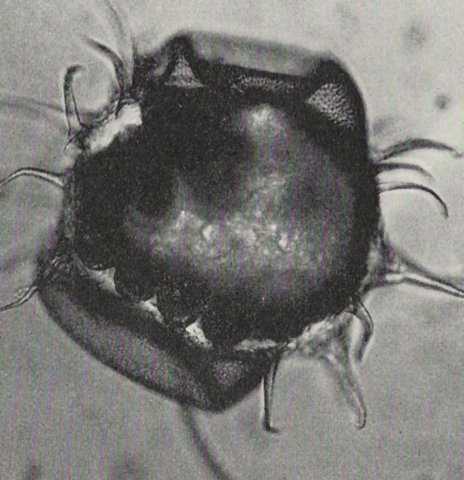
Fig.4. Egg of Orchesella cincta After Bretfeld in Schaller, 1970:51
In some cases, the eggs which are at first apparently smooth, after a few
days will be found to be covered with long hairs (Lubbock, 1873:83).
The after the chorion split freed vitelline membrane surface carries
in many species (such as in Orchesellinae, Heteromurinae and Tomoceridae)
numerous processes (fig.2 right, fig.4, fig.4aa, fig.4ab),
in other species (such as in Onychiuridae) flattened nipples
(fig.2 middle)
(Kühnelt, 1961:162; Schaller, 1970:51).
These structures enable the egg to adhere firmly to the substrate
on which it has been deposited
(Kühnelt, 1961:162)
(protection against flotation due to in the soil penetrating rain water)
(Schaller, 1970:51).
Fig.4a. Folsomia candida Egg in anaphase
Chorion is split in two polar caps
After Gao et al. 2006 Fig.3.5
A schematic animation of the formation of the polar caps of the egg of
Folsomia candida WILLEM, 1902
based on observations of a specimen culture kept from 1994 to 1996
(Janssens, 1994:6-10; Janssens, 1996:120-125):
fig.5 shows three orthogonal views at the same time:
upper left: frontal view,
upper right: lateral view, and
lower left: dorsal view.
Due to the swelling of the egg, the chorion bursts at the equator of the egg.
The two halves of the chorion
are pushed towards the poles of the egg (Fig.4b, 4ba),
while the originally spherical egg
evolves into a laterally flattened ellipsoid shape.
The chorion halves fold in the shape of
a craterlike depression at the poles (Fig.4bb),
forming in such a way two polar caps.
The polar caps are eventually 'ejected' from the egg surface (not shown).
Fig.4d. Cryptopygus antarctica Chorion is split in two polar caps
After Schaller & Kopeszki 1991 Abb.4
In Cryptopygus antarcticus the orangereddish spherical eggs are
deposited in packets (Fig.4c).
The diameter of the egg is 0.2 mm.
The number of eggs per packet is 6 to 9.
The female keeps the eggs in a packet together by circumventing the eggs
with sticky threads from the genital opening.
The egg packet survives a flood.
Egg packets are often clustered to form a giant packet.
At 0-2 degrees Celsius the eggs hatch after 144 days.
At 7-9 degrees Celsius the embryonal development takes 28 to 34 days.
The chorion splits after 8 days (Fig.4d). The egg deforms to a more oval shape:
the diameter increases to 0.23 mm while the egg thickness reduces to 0.18 mm.
The egg colour underneath the chorion is yellowish.
(Schaller & Kopeszki 1991:222-223).
In Sminthurides aquaticus, at the end of the embryonic development, the
eggshell is completely transparant revealing the fullgrown embryo ready to hatch.
(Fig.3b,3bb)
In Folsomia candida, at the end of the embryonic development, the
diameter of the egg reaches 0.180-0.185 mm.
(Gao, Bu, Luan & Yin, 2006:519).
Freshly deposited eggs are orange-brown. The egg swells during embryonic
development, the chorion bursts and forms two polar caps.
The newly hatched instar is about 0.35 mm in length.
Newly hatched young of Isotoma viridis measure 0.57-0.63 mm in
length and 0.15 mm in head width. A red pigment, possibly carotenoid is
present in the body fluid of this species, giving the young a pale red
colouration (Milne, 1960). The surface pigmentation only becoming apparent
after the first ecdysis (Milne 1960).
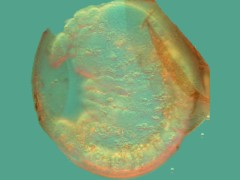
A freshly hatched unidentified Isotomidae is completely exhausted.
Acknowledgements
The author wishes to thank, in alphabetical order,
Ernest C. Bernard,
Nathan Paiva Brito,
P.M. Chamberlain,
Hsin-Ju Cheng,
Marie-Louise Huskens,
Charlene Janion,
Terry Lynch,
D. Sikes,
Jan van Duinen,
Andrew Robertson,
Domingo Zungri
for the usage of their images as illustrations.