
6. Literatuur
Populieren zijn bomen die af en toe in de loofbossen van de gematigde zone van het noordelijk halfrond voorkomen (Engelen, 1998). Ze zijn gebonden aan vochtige gebieden en kunnen zelfs groeien in gebieden die regelmatig overstroomd worden door beken of rivieren. Er komt in België van nature één soort populier voor. Dit is de ratelpopulier of esp (Populus tremula L.). Verwilderde soorten in België zijn onder andere de witte abeel (Populus alba L.), de grauwe abeel (Populus canescens (Ait.)Smith) en de zwarte populier (Populus nigra L.) (Lambinon, De Langhe, Delvosalle & Duvigneaud, 1998). Wegens commerciële redenen zijn er echter zeer veel klonen van kruisingen ontwikkeld die algemeen aangeduid worden onder de naam Populus x euramericana GUINNIER (Engelen, 1998).
Ongeveer de helft van alle populieren in Vlaanderen groeit in de zand- en de zandleemstreek, nog eens een kwart in de leemstreek en ongeveer zestien procent in de Kempen (Engelen, 1998).
|
|
1.1.2 Leeftijdsevolutie
Uit een studie uit 1995, uitgevoerd door het Laboratorium voor Bosbouw van de Rijksuniversiteit Gent en het Koninklijk Instituut voor het duurzame beheer van de natuurlijke rijkdommen en de bevordering van schone technologie (De Keersmaeker & Muys, 1995), is gebleken dat een populierenaanplant op dezelfde wijze zal evolueren als een ander bos. De enige reden dat populierenbossen bijna nooit een climaxvegetatie hebben, is dat deze bossen gerooid worden voor ze oud genoeg zijn om een dergelijke vegetatie te herbergen. De aanwezigheid van ruderale planten zoals grote brandnetel (Urtica dioica L.) is enkel te wijten aan de jonge leeftijd van de bossen en aan een dikwijls voorkomende tijdelijke voedselaanrijking. Deze aanrijking ontstaat doordat populieren vaak op drassige gronden geplant worden. Om goed te groeien en voor gemakkelijk onderhoud wordt vaak sterk gedraineerd. Ook de populieren zelf zuigen redelijk wat water weg. Hierdoor kan de voorraad organisch materiaal in de bodem op korte tijd verteerd worden, nadat er terug zuurstof in de bodem gekomen is. Dit heeft een tijdelijke opstoot in het stikstof- en fosforgehalte tot gevolg, waar vooral de grote brandnetel (Urtica dioica L.) van profiteert.
1.1.3 Bostypes
In het kader van het project is er een vegetatiestudie verricht op 189 verschillende plots. De plots van deze bossen liggen over heel Vlaanderen verspreid. Hierbij moet wel vermeld worden dat de provincie West-Vlaanderen zeer weinig plots heeft, omdat het bosareaal in deze provincie beperkt is. Twee van de plots hadden geen enkele andere soort bomen of struiken behalve de populier zelf en zijn buiten de verwerking gelaten (Verstraeten, 2001).
Op de 187 overige plots werd een vegetatieopname uitgevoerd door het Instituut voor Bosbouw en Wildbeheer. Enkel de planten in de kruidlaag werden hierin opgenomen, zaailingen van bomen niet. De gegevens zijn verwerkt met het classificatieprogramma TWINSPAN. Dit programma heeft zeven hoofdtypes van begroeiing herkend. Deze zijn:
- graslandtype: hier groeien voornamelijk grassen en algemene planten zoals witte klaver (Trifolium repens L.) en ridderzuring (Rumex obtusifolius L.). Grote brandnetel (Urtica dioica L.) komt voor, maar met gemiddeld minder dan 5% bedekking;
- soortenarm brandneteltype: grote brandnetel (Urtica dioica L.) heeft een gemiddelde bedekking van meer dan 58 % en alleen hondsdraf (Glechoma hederacea L.) en kleefkruid (Galium aparine L.) komen nog geregeld voor;
- speenkruid-brandneteltype: hier staat gemiddeld meer dan 55 % grote brandnetel (Urtica dioica L.), maar ook bijna altijd speenkruid (Ranunculus ficaria L.) met eveneens een vrij hoge bedekkingsgraad. Andere soorten die hier voorkomen zijn zich snel verspreidende soorten zoals bijvoorbeeld hondsdraf (Glechoma hederacea L.), kleefkruid (Galium aparine L.), geel nagelkruid (Geum urbanum L.), engelwortel (Angelica sylvestris L.) en bosandoorn (Stachys sylvatica L.);
- ruderaal brandneteltype: dit type heeft meer dan 52 % grote brandnetel (Urtica dioica L.) en af en toe ook speenkruid (Ranunculus ficaria L.). Verder komen gelijkaardige soorten voor als bij het speenkruid-brandneteltype, alsook enkele ruderale plantensoorten zoals smeerwortel (Symphytum officinale L.), moesdistel (Cirsium oleraceum (L.) Scop.), akkerdistel (Cirsium arvense (L.) Scop.), en gewone berenklauw (Heracleum sphondylium L.);
- braam-brandneteltype: grote brandnetel (Urtica dioica L.) en braam (Rubus sp. L.) hebben hier beide een bedekking die gemiddeld tussen 12,5 en 50 % ligt. Verder staan hier gelijkaardige planten als in de vorige twee types. We vermelden ook nog witbol (Holcus sp. L.) en pitrus (Juncus effusus L.);
- verzadigd type: grote brandnetel (Urtica dioica L.) haalt toch geregeld nog een gemiddelde boven 5 % bedekking en grassen zijn bijna volledig afwezig, maar er zijn wel zeer veel soorten echte bosplanten. De standaardsoorten zoals kleefkruid (Galium aparine L.) en hondsdraf (Glechoma hederacea L.) komen voor, maar ook bijvoorbeeld bosanemoon (Anemone nemorosa L.), gewone salomonszegel (Polygonatum multiflorum (L.) All.), gevlekte aronskelk (Arum maculatum L.) en muskuskruid (Adoxa moschatellina L.);
- zeer soortenarm type: in dit type bossen, waar er maar vier van tussen de plots zaten, staan bijna geen andere planten. Zelfs grote brandnetel (Urtica dioica L.) ontbreekt hier vaak. De bomen nemen vaak zeer veel licht weg doordat de kronen dicht op elkaar aansluiten. Eén van de bossen had bijvoorbeeld een combinatie van beuk (Fagus sylvatica L.) met populier, waarbij de beuken het meeste licht wegnamen.
Verder kunnen we stellen dat de types veelal leeftijdsgebonden zijn. Ze zijn ook afhankelijk van de voorgeschiedenis van het bos. Het aantal soorten planten in oude bossen neemt in aantal en bedekkingsgraad enkel maar toe met de leeftijd. Enkele plantensoorten, zoals kruipend zenegroen (Ajuga reptans L.) en breedbladige wespenorchis (Epipactis helleborine (L.) Crantz), zijn typisch voor jong bos en deze soorten zullen dan ook verdwijnen naarmate het bos veroudert. Andere effecten die in acht moeten genomen worden behalve leeftijd zijn bijvoorbeeld begrazing en isolatie- en randeffecten. Door begrazing zal een bos meestal tot het graslandtype behoren.
Ook de struiklaag is van belang, omdat deze door het wegnemen van licht andere planten geen kans geeft. Grote brandnetel bijvoorbeeld krijgt problemen bij te veel schaduw. Er zijn volgens TWINSPAN vijf types struiklaag in de onderzochte bossen:
- type 1: hazelaar met soms rode kornoelje en wilde kamperfoelie;
- type 2: vlier;
- type 3: zwarte els en soms hop;
- type 4: zomereik en soms braam;
- type 5: boswilg en soms ruwe berk.
De overeenkomst tussen de begroeiingstypes en de struiklaagtypes vindt u in onderstaande tabel (Verstraeten, 2001).
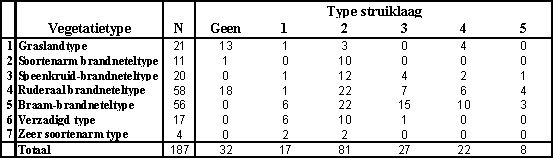
Opvallend is de overeenkomst tussen soortenarm brandneteltype en de dominantie van vlier. In de oudere types, verzadigd en zeer soortenarm type, krijgen we vaak vlier en hazelaar, die typisch is voor oude bossen. In de twee jongste types, grasland- en ruderaal brandneteltype, hebben we de enige plots zonder struiklaag. Dit kan zijn omdat de struiken gewoon nog niet de kans gehad hebben om deze jonge bossen te koloniseren.
In de buurlanden wordt populier verder ook wel eens gebruikt als pioniervegetatie, om zo sneller tot een begroeid bos te komen. Mogelijk kunnen hierna de populieren gedeeltelijk of volledig vervangen worden door inheemse boomsoorten. Of een dergelijk bos verschilt in vegetatie ten opzichte van een bos dat op natuurlijke wijze ontstaat, moet nog onderzocht worden.
|
 |
De Collembola of springstaarten zijn primitieve dieren, waarvan de juiste positie binnen het dierenrijk nog ter discussie staat. Vroeger werden ze door de aanwezigheid van zes poten tot de insecten gerekend en dan meer bepaald als een orde binnen de oerinsecten of Apterygota. In deze groep vonden we ook de ordes van de Protura, de Diplura en de Thysanura, waartoe het bekende zilvervisje, Lepisma saccharina, behoort. De groep van de Apterygota wordt niet meer gebruikt in de cladistische systematiek, bij welke evolutieve verbanden en onderlinge verwantschap primeren, gezien hij een samenraapsel is van een aantal parafyletische groepen. Tegenwoordig worden Collembola, Protura, Diplura en Thysanura als vier aparte klasses gezien (Janssens, 2002).
Er zijn momenteel ongeveer 7000 gekende soorten Collembola, in grootte variërend van 0,2 tot 10mm, waarvan er zelfs op Antarctica voorkomen. Dit maakt de springstaarten tot de meest verspreide groep zespotigen. Het oudste, gekende fossiel van een springstaart stamt uit het Devoon, wat springstaarten zowat de oudste groep van de Hexapoda maakt en tevens ook één van de eerste groepen landdieren. Het is al even merkwaardig dat er nu nog soorten Collembola bestaan die amper verschillen van het fossiele exemplaar, waaruit niet alleen de hoge primitiviteit van sommige recente soorten blijkt, maar ook het enorme aanpassingsvermogen van deze diergroep (Janssens, 2002).
Het lichaam van de Collembola bestaat uit drie delen: de kop, het borststuk of thorax en het achterlijf of abdomen. Collembola bezitten zoals alle geleedpotigen segmenten. De thorax bezit drie segmenten, met elk één potenpaar. Het abdomen bevat zes segmenten, zelfs in embryonale fase, terwijl de insecten in ieder geval tijdens dit stadium altijd elf segmenten hebben. De laatste twee segmenten zijn meestal vrij klein ten opzichte van de vier andere. De ogen van springstaarten zijn niet gefacetteerd, zoals bij de insecten, maar bestaan uit een pigmentvlek met daarop nul tot acht ocelli. Verder bezitten Collembola een orgaan dat PAO of post-antennaal orgaan genoemd wordt. Het is zeer variabel van vorm, maar ligt altijd achter de antennes. Het is een chemoreceptor, die waarschijnlijk ook een hygrometerfunctie heeft. De monddelen zijn moeilijk herkenbaar, gezien ze in het kopkapsel verzonken liggen. De poten zijn geleed, maar in vergelijking met insecten primitief van bouw. Er kunnen een aantal leden onderscheiden worden. Dit zijn vanaf het lichaam af: subcoxa, coxa, trochanter, femur, tibiotarsus en pretarsus. De klauw met het empodium zit vast aan de pretarsus. Deze klauw is zeer belangrijk als determinatiekenmerk (Joosse-Van Damme, 1963).
Collembola bezitten een aantal vreemde aanhangsels aan het lichaam. Op het eerste abdominale segment is er de collofoor of ventrale tubus, een dunwandige uitstulpbare buis met twee ventrikels of zakjes aan het uiteinde. De functie van de ventrale tubus is nog niet helemaal duidelijk, maar waarschijnlijk doet het dienst als een vasthechtingsorgaan bij het voortbewegen op gladde oppervlakken, als orgaan voor wateruitwisseling, of zelfs voor uitwisseling van chemicaliën met de omgeving. Op het derde abdominale segment is er het retinaculum, een tangachtig apparaat dat dienst doet voor de in rust zijnde springvork. Deze springvork of furca, wellicht het duidelijkste kenmerk van de meeste Collembola, staat op het vierde abdominaal segment. Hij bestaat uit een ongepaard manubrium met daarop twee dentes, die elk in een mucro eindigen. Deze mucro is ook van groot belang bij de determinaties. Een springstaart kan bij een gevarensituatie de furca achterwaarts uitklappen, waardoor het dier zonder duidelijke richting weggekatapulteerd wordt (Janssens, 2002).
Springstaarten hebben gescheiden geslachten, maar de genitaliën zijn inwendig, waardoor onderscheid tussen de seksen zeer moeilijk vast te stellen is, behalve bij een paar genera, zoals Orchesella, waarbij een aantal soorten een duidelijk geslachtsdimorfisme vertoont (Joosse-Van Damme, 1963).
Zoals bij alle Arthropoda groeit de huid niet mee en zijn er dus verschillende vervellingen in het leven van een springstaart, die zelfs verdergaan nadat het dier volwassen is. Op deze manier worden ook afvalstoffen uitgescheiden. Deze stoffen worden in het darmepitheel opgeslagen door het ontbreken van buisjes van Malpighi. Het darmepitheel regenereert mee bij de vervelling (Joosse-Van Damme, 1963).
Binnen de Collembola komen drie ordes voor: de Poduromorpha, de Entomobryomorpha en de Symphypleona. De beschrijving van deze ordes is gebaseerd op de tekst van Joosse-Van Damme(1963), aangevuld met mondeling verkregen informatie van de heer Frans Janssens en met eigen observaties.
De Poduromorpha zijn gedrongen van vorm, vaak met uitstulpende segmenten. De antennes zijn vrij kort en ook verdikt. Ze zijn homogeen grijs of blauwgrijs gekleurd of missen elke pigmentatie. De furca is meestal sterk gereduceerd of zelfs volledig afwezig. Het huidoppervlak van deze groep is dikwijls onregelmatig. De familie van de Onychiuridae bezit bovendien ook nog eens pseudocelli op het lichaam. Deze orgaantjes hebben een functie bij hun verdediging. Collembola bezitten de mogelijkheid om bij groot gevaar aan reflexbloeden of exudatie te doen. Hierbij wordt hemolymfe of bloedvloeistof op speciale plaatsen naar buiten geperst. Waar dit proces bij de meeste Collembola op willekeurige plaatsen gebeurt, zijn deze plaatsen bij de Onychiuridae de pseudocelli. Verder bezitten de Poduromorpha ogen die een reductie vertonen van zes tot nul ommatidiën.
|
 |
|
 |
 |
|
De meeste Collembola planten zich voort door middel van spermatoforen. De mannetjes zetten deze achteloos en veelvuldig af op alle mogelijke plaatsen, zonder zich blijkbaar door de vrouwtjes te laten beďnvloeden. Deze moeten de spermatoforen dan vinden, vermoedelijk door op de geur af te gaan. Oude spermatoforen worden veelvuldig geconsumeerd door zowel mannetjes als vrouwtjes, wat ervoor zorgt dat er continu verse spermatoforen voorradig zijn (Joosse-Van Damme, 1963).
Bij de Symphypleona merken we echter een ander gedrag op. Hier zijn er soorten waar de mannetjes complexe antennes bezitten, waarmee ze die van het vrouwtje kunnen vastgrijpen. De mannetjes kunnen dan dagenlang het vrouwtje ronddragen. Bij één geslacht, Bourletiella, zet het mannetje de spermatofoor op de grond af. Daarna wordt de spermatofoor over de voorpoten uitgesmeerd. De voorpoten worden naar de mond gebracht en met behulp van de monddelen worden de spermacellen in de vrouwelijke geslachtsopening gebracht (Joosse-Van Damme, 1963).
Collembola vertonen protomorfose als metamorfosetype. Hierbij ontstaan er geen nieuwe segmenten bij de vervellingen na het embryonaal stadium en gaan de vervellingen door, ook nadat het dier volwassen geworden is. Dit zijn de zogenaamde imaginaalvervellingen (Joosse-Van Damme, 1963).
Collembola worden onder laboratoriumomstandigheden 5 tot 18 maanden oud, met één jaar als standaardleeftijd. Dit hangt van soort tot soort af. Jonge Collembola zijn volledig wit en krijgen met elke vervelling meer pigment. Ze vervellen tot 50 keer in hun leven. De chitinehuid zal bij oudere dieren alsmaar sterker geel kleuren, door opslag van onafbreekbare excretieproducten. Deze kunnen niet langs andere weg afgevoerd worden door het ontbreken van buizen van Malpighi (Joosse-Van Damme, 1963).
De frequentie van vervellen is variabel en gebonden aan onder andere temperatuur. Lindenmann vond bij Orchesella een variatie van 2 tot 29 dagen bij de exemplaren van éénzelfde legsel. Na tien tot vijftien vervellingen zijn de dieren geslachtsrijp en na een twintigtal vervellingen is er geen toename in grootte meer. In de winter treft men hoofdzakelijk adulte exemplaren van alle springstaarten aan en zelfs in het labo bij hogere temperaturen gaan deze niet of nauwelijks over tot voortplanting. In de rest van het jaar zijn de populaties gewoonlijk gemengd, met soortafhankelijke pieken in het voorkomen van juvenielen (Joosse-Van Damme, 1963).
Collembola zitten meestal in de grond of in humus en verkiezen een vochtige of natte omgeving, hoewel sommige soorten eerder op schors of bloemen gaan zitten. Een aantal typische habitats zijn mos, grotten, mieren- en termietennesten en zelfs wateroppervlakken en sneeuw. Springstaarten zijn zeer belangrijk in bodemecosystemen en nemen in een gemiddeld bos toch een paar procent van de biomassa in. De meeste Collembola voeden zich met micro-organismen in de bodem, zo zijn er zeer veel schimmeleters, of met organisch materiaal dat ze zo verder composteren. Ook eencellige algen behoren vaak tot de voeding. Er zijn ook een aantal predatoren gekend onder de springstaarten. Collembola zorgen verder ook nog voor de verspreiding van schimmels en bacteriën doordat deze aan de dieren blijven hangen als ze verder kruipen. Springstaarten zijn echter wel te klein om de poriënstructuur van de bodem te wijzigen en ze zijn dus niet in staat om compacte bodems te koloniseren (Joosse-Van Damme, 1963).
Collembola zijn zowel indicatoren van bodemeigenschappen als van menselijke verstoring. Elke soort springstaart reageert anders op chemische bodemparameters. Sommige soorten reageren niet bij verandering van deze parameters, maar de meeste soorten worden door een drietal parameters beďnvloed. Deze parameters verschillen onderling voor de verschillende soorten. Door bij een onderzoek van een bepaald gebied de resultaten voor meerdere soorten te combineren kan men een volledig overzicht krijgen van de bodemtoestand in dit gebied. Sommige soorten kunnen zelfs voor zes eigenschappen als indicator gebruikt worden, namelijk voor pH, baseverzadigingsgraad, Ca, Mg, Mn en K. Deze soorten kunnen op zich als indicator dienen, gezien zij alle verschillend op de zes parameters reageren. Collembola kunnen ook als indicator dienen voor het humustype en voor de heterogeniteit van de bodem (Jacobs, 1997).
Menselijke verstoring heeft invloed op het bodemecosysteem en hiervoor zijn Collembola ook geschikte indicatoren. Bodemverstoring kan zowel fysisch als chemisch zijn. Collembola hebben wel beperkingen als indicator hier, gezien ze gevoelig zijn voor verandering van de pH, zwaveldioxide-immissie en bodemverdichting. Daarentegen zijn ze ongevoelig voor kopervervuiling in wijngaarden (Jacobs, 1997).
Collembola zijn zeer sterk vochtgebonden. Het vochtgehalte in de bodem is dan ook de belangrijkste bepalende factor voor het aantal springstaartexemplaren in een gebied. Zo zien we dat het aantal dieren in een vochtiger habitat steeds hoger ligt dan in een droger habitat van hetzelfde gebied. Veranderingen in de populatie zijn meestal gebonden aan veranderingen van het bodemvochtgehalte. Het aantal soorten wordt hierdoor echter minder beďnvloed (Kaczmarek, 1995).
Ook de verteerbaarheid van het bladstrooisel speelt indirect een rol. Bladstrooisel van boomsoorten die snel afgebroken worden en dus kwalitatief goede humus leveren, heeft een hogere temperatuur en een lagere vochtigheid dan dat van boomsoorten waar het afbraakproces trager verloopt en de humus dus een lagere kwaliteit heeft. Strooisel van deze traag ontbindende bladsoorten bevat veel meer Collembola dan bladstrooisel van de gemakkelijker af te breken soorten. Hierin speelt de hogere vochtigheid van het laag kwalitatieve strooisel een belangrijke rol, waardoor we terug uitkomen bij het belang van het bodemvocht (Badejo, Nathaniel & Tian, 1998).
Ook bij een vergelijking in Italië tussen een eikenbos met mullhumus, een eikenbos met morhumus en twee naaldbossen met ruwe humus komt dit verschijnsel terug. De ruwere humustypes bevatten veel grotere aantallen springstaarten. Wel van belang om hier te vermelden is dat de mullhumus de hoogste soortendiversiteit heeft en de ruwe humus de laagste. Er komen in de naaldbossen wel een paar soorten voor die niet in de eikenbossen terug te vinden zijn (Kopeszki & Meyer, 1996). Hierop aansluitend is een ander onderzoek van belang, namelijk uit de Franse Pyreneeën. Hier werden natuurlijke en op landbouwgrond aangeplante berkenbossen vergeleken met naaldboomaanplantingen, grenzend aan de besproken berkenbossen. Het aantal soorten in de naaldbossen lag ook hier veel lager dan in de berkenbossen, maar specifieke studie naar de endemische soorten, toch wel de belangrijkste component van de fauna op vlak van natuurbehoud, gaf een drastische vermindering in de diversiteit weer (Deharveng, 1996). Het aanbrengen van snoeisel in nieuwe bossen om sneller een humuslaag te verkrijgen kan dus best gebeuren met inheemse boomsoorten.
In Collembolapopulaties zijn seizoenseffecten waar te nemen. Deze verschillen echter bij naald- en loofbossen. Hieruit volgt dat ook de aanwezigheid van voedsel een effect op de populatie heeft. Bij naaldbossen is er immers een constante aanvoer van voedsel, terwijl er bij loofbossen een plotse grote aanvoer in de herfst is. De constante voedselvoorziening kan mee verklaren waarom in naaldbossen grotere aantallen springstaarten leven (Kaczmarek, 1977).
Collembola kunnen gebruikt worden om de kwaliteit van hun biotoop te onderzoeken. Standaard kunnen we stellen dat in een natuurlijk, onverstoord biotoop, weinig micro-artropoda, in dit geval springstaarten, een afwijkende levenswijze zullen hebben. Hoe meer verstoring er is, gaande van een eenmalige storting van wat tuinafval, over begrazing en intensief maaibeheer, tot vervuiling met zware metalen en persistente pesticiden, hoe meer afwijkende gedragingen we kunnen waarnemen. Deze afwijkende gedragsverschijnselen blijken constant genoeg te zijn om vergelijkingen te maken tussen verschillende gebieden en landen (Siepel, 1995).
Zo leiden onregelmatige verstoringen tot een stijging van foresie tactieken. Foresie is het vasthechten van een dier op een ander, zonder het schade te berokkenen. Het dier dat deze tactiek gebruikt stopt met zich te voeden en met groeien. Zo is er een veel grotere verspreiding mogelijk en kan de populatie beter overleven dan in een deel van het gebied waar plotseling een verstoring optreedt (Siepel, 1995).
Wanneer de verstoringen echter met een vaste tussenperiode voorkomen, zien we een verhoogde vertegenwoordiging van synchronisatietactieken. Hierbij passen de dieren zich zodanig aan aan de verstoringen dat hun cyclus of een deel ervan juist tussen twee verstoringen valt (Siepel, 1995).
Een laatste duidelijke aanpassing vinden we bij een vervuiling met zware metalen of andere polluenten. Hier vinden we een sterke stijging in thelytoky. Dit is een parthenogentische voortplanting waarbij de nakomelingen allemaal vrouwelijk zijn. Dit wordt veroorzaakt door het feit dat de weinige dieren die in eerste instantie resistent zijn tegen de vervuiling de populatie sneller weer moeten laten herstellen. Bovendien kan de resistentie bij geslachtelijke voortplanting snel weer verloren gaan, waardoor een ongeslachtelijke voortplanting interessanter is (Siepel, 1995).
Als we de evolutie naar een typische bosfauna van eender welke groep organismen bekijken, moeten we ten eerste een onderscheid maken tussen nieuw beboste gronden en gronden die herbebost worden kort na een kaalslag. Als een jong bos begint te groeien zal het een schermfunctie krijgen waardoor de sterke schommelingen in temperatuur en bodemvocht getemperd zullen worden. Er ontstaat een typisch bosklimaat (Verstraeten, 1999).
Naar gelang van de diergroep duurt het iets meer dan een decennium tot zelfs een paar decennia eer er typische bossoorten van de groep in kwestie beginnen te verschijnen. Indien een gebied herbeplant wordt kort na een kaalkap, zijn deze periodes merkelijk kleiner. Als regel kan worden aangenomen dat bij bebossing van landbouwgronden de macrofaunagemeenschap zich pas na 60 tot 100 jaar kan ontwikkelen tot een volwaardige bosgemeenschap. Na kaalkap en herbeplanting duurt het 25 tot 30 jaar om dit stadium te bereiken. Bij een nieuwe aanplant domineren na 25 tot 35 jaar nog steeds typische landbouwgrondsoorten (Verstraeten, 1999).
Bij verschillende boomsoorten gebeurt deze evolutie met een verschillende snelheid. Bij een populierenaanplanting kan door de snelle groei reeds na een decennium een bosklimaat ontwikkeld zijn (Verstraeten, 1999).
Ook moet vermeld worden dat de snelheid van kolonisatie van een landbouwgrond manueel kan versneld worden, althans toch voor de planten. Door het inzaaien van vaste plantensoorten kan de kruidlaag sneller een boskarakter krijgen. Het inzaaien van eenjarigen, welke vanzelf als eerste de landbouwgrond zouden innemen, vertraagt echter de kolonisatie niet (Wilcox, 1998). Gezien de plantendiversiteit zo sneller opgedreven kan worden, is het mogelijk dat door aanvoer van boshumus in een nieuwe aanplanting ook de invertebratendiversiteit sneller deze van een bos kan bereiken. Dit moet echter nog onderzocht worden.
De evolutie van de bodeminvertebraten hangt ook af van het bestaande humustype voor bebossing. Als men vertrekt van een weiland met een goede mullhumus en het bladstrooisel van de aanplant is van goede kwaliteit, dan blijft deze goede humus behouden. Indien het bladstrooisel echter van slechte kwaliteit is, krijgen we na verloop van tijd een moderhumus met een veel kleinere biomassa aan bijvoorbeeld regenwormen. Ook de pH en de baseverzadigingsgraad zullen zo verschillend evolueren bij bebossing van weiland (Jacobs, 1997).
Bij kaalkap en herbebossing is het van belang te weten dat de bodemeigenschappen tot tien jaar na de kaalkap weinig veranderen. Kleine verschillen komen voor in temperatuur en bodemvocht, alsook op vlak van bodemdichtheid en een aantal chemische parameters. Verder is de afbraaksnelheid van humus wat hoger in een kapvlakte dan in een bos. Maar als er direct na een kaalkap herbebost wordt, zal het bodemleven hier op termijn niet zo veel invloed van ondervinden (Houston, Visser & Lautenschlager, 1998).