2. Inleiding
1. Systematiek en fylogenie der klasse Collembola
Collembola worden dezer dagen beschouwd als zijnde een Klasse van het Fylum Arthropoda, doch hun exacte taxonomische positie is nog steeds het onderwerp van enig debat.
De Klasse Collembola, als monofyletische groep, wordt gekenmerkt door een ventrale tubus en een springorgaan, de furca, sommigen vertonen een secundaire reductie van het laatstgenoemde. De antennen bestaan uit vier segmenten, welke soms onderverdeeld in subsegmenten, en er zijn zes abdominale segmenten, vergroeid bij Neelipleona en Symphypleona.
Tot de Poduroidae behoren die families, waarbij het voorste segment van de thorax, de prothorax, dorsaal duidelijk zichtbaar en altijd op zijn minst enkele dorsale setae draagt. De structuur van de monddelen is belangrijk voor het scheiden in de families. Poduroidae zijn geneigd zich traag voort te bewegen en leven meestal in de grond, onder stenen, schors, of afgevallen bladeren. Sommigen hebben de mogelijkheid om te springen verloren. De Entmobryoidae daarentegen zijn actievere springstaarten met een sterk gereduceerde prothorax, welke nooit dorsale setae draagt, met een grotere bekwaamheid om te springen. De Neelipleona omvat een enkele familie Neelidae van kleine, blinde bodembewoners, welke globulair in vorm zijn. De Symphypleona, evenzo rondvormig, met gefuseerde, abdominale segmenten bezitten de mogelijkheid tot springen en zijn algemeen in bovengrondse vegetatie.
De eerder conventionele, ordinale fylogenetische verwantschap, voorgesteld door Janssens (2001), is gebaseerd op een lineaire fylogenie van de progressieve, toenemende functie van de furca. Er wordt verondersteld dat bij de protocollembola de furca niet goed ontwikkeld is. Initieel is zij klein: een furcula. De Poduromorpha is de oudste groep in bezit van zo een kleine furca. De springvork reduceert zelfs verder in plaats van te ontwikkelen. De ‘hogere’ Collembola, daarentegen, specialiseren zich in een goed ontwikkelde furca. De Metaxypleona is de eerste groep van hogere collembolen met een grote furca. Podura aquatica is de enige overlevende van deze oude stam van springstaarten.
De grotere springvork heeft zijn uitwerking op de lichaamsvorm: het wordt dorso-ventraal gebogen te wijten aan de grote antero-posterieure spieren, die de furca bedienen. De Neelipleona worden hier beschouwd als de oudste zijtak van de ‘nieuwe’ Collembola, gekarakteriseerd door een gereduceerde prothorax. In de hoofdstam ontwikkelt het abdomen zich verder als gevolg van de grotere furcaspieren, resulterend in een dorsaal gebogen lichaam. Bij de Entomobryomorpha is de curvatuur relatief zwak, een komma-vormige laterale habitus als gevolg. De furca kan gereduceerd zijn of zelfs afwezig, de kromming van het lichaam nog verkleinend. De lichaamskromming bereikt haar hoogtepunt bij de Symphypleona: het laatste thoraxsegmenten en de eerste abdominale segmenten zijn opwaarts gebogen, zodanig dat ze een inverse ‘U’ vormen, effectief de lichaamslengte verkortend.
Tabel 1: Classificatie van de Collembola (Janssens, 2001)
2.Morfologie van de Collembola
2.1. Algemene lichaamsbouw
Springstaarten zijn kleine, primitieve, vleugelloze Hexapoden. Hun lichaam is duidelijk onderverdeeld in een kop, een thorax met drie segmneten en een abdomen met zes segmenten. Het lichaam is bedekt met haren (setae) of schubben ( gemodificeerde haren) van verschillende vorm. De schikking en het aantal haren of schubben is meestal karakteristiek op soortniveau.
De antennen, opgebouwd uit vier basissegmenten, rijkelijk bezet met sensorische organen (Eisenbeis &Wichard, 1987; Nayrolles, 1991), zijn ingeplant op een verheven deel van de kop, nl. de antennale basis. Sommige soorten bezitten antennen, die nog eens onderverdeeld zijn in subsegmenten en aldus met een grotere flexibiliteit. Het verlies van de antennes komt veelvuldig voor bij de Collembola, het is te wijten aan aanvallen van roofzuchtige kevers, na verschillende vervellingen kunnen de antennen volledig terug gegroeid zijn (Ernsting & Fokkema, 1983). Tussen de ogen en de antennale basis vindt men meestal het postantennaal orgaan (Figuur 5) terug, bestaande uit een enkel ovoïd lichaam ofwel uit meerdere lichamen van variërende vorm en samenstelling (Fjellberg, 1980). Dit sensorisch orgaan zou een olfactorische functie hebben.
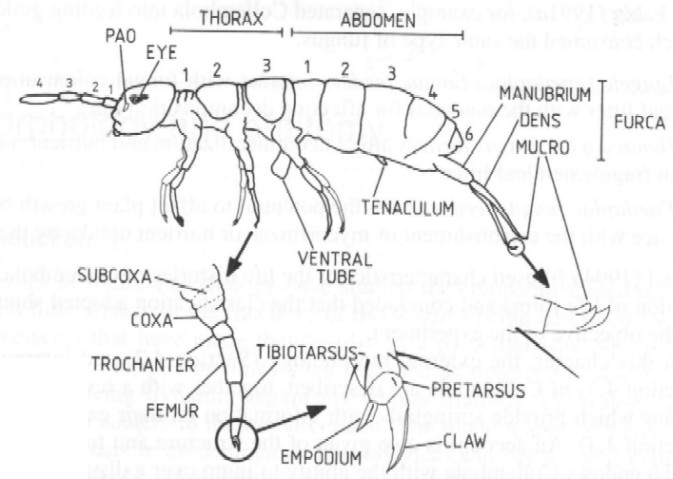
De thoracale segmenten dragen elk een paar poten. Iedere poot bestaat uit een subcoxa, een coxa, een trochanter, een femur, een tibiotarsus en een pretarsus met een klauw en een empodium (Figuur 1). Bij bepaalde soorten zijn de klauwtjes aangepast om makkelijker op natte oppervlakken te lopen (Deharveng & Gouze, 1986; Fjellberg, 1991; Palévody, 1965). Bij andere soorten draagt de tibiotarsus één of meerder lange haren met kleverige uiteinden, zodat het dier kan blijven kleven aan vlakke oppervlakken (Blottner & Eisenbeis, 1984).
Het eerste abdominale segment van alle Collembola draagt een ventrale tubus, welke een belangrijke rol speelt bij de vloeistof- en elektrolietenbalans. Het is een enigszins uitstulpbare, zeer dunwandige blaas, die soms gebruikt wordt als zuignap bij de voortbeweging over gladde oppervlakken (cfr. naam, "colle" = lijm en "embolon" = zuignap of pen). Het derde segment van het abdomen draagt een tenaculum of retinaculum, dat dienst doet als haak voor het springorgaan of furca, die op het vierde abdominale segment is ingeplant. De structuur en functie van de furca worden later nog besproken (zie 2.5.). Aan de ventrale zijde van het vijfde abdominaal segment zit de onopvallende mannelijke of vrouwelijke genitale opening. Het spijsverteringsstelsel eindigt via de anus op het zesde abdominaal segment, dat tevens drie zakjes draagt, waarvan de functie nog niet gekend is (Leinaas, 1988).
De plaats waar de springstaarten leven, is kenmerkend voor hun lichaamsbouw. Op morfologisch-ecologische gronden kan men zo drie verschillende vormen onderscheiden:
2.2. De monddelen
De Collembola zijn entognaat: hun monddelen ontwikkelen zich tijdens de embryogenesis in de buccale holte in de kop (Uemiya & Ando, 1987). Er zijn vijf basisonderdelen van de mond,
zijnde een labrum (bovenlip), een paar mandibulae (bovenkaken), een paar maxillae (onderkaken), een hypopharynx (onpare tong) en een labium (onderlip). De maxillaire palpen of kaaktasters vindt men beiderzijds van dit complex. De monddelen zijn dorsaal ingesloten door het labrum, ventraal door het labium en lateraal door de pleurale plooien van de cuticula (Figuur 2). De toppen van de mandibels zijn sterk getand, waarmee voedsel van het substraat geschraapt kan worden (Christian, 1989).
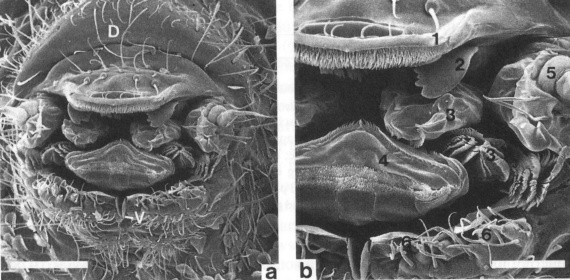
Figuur 2: Opname van de monddelen van Pogonognathellus flavescens(Tomoceridae) met een elektronenmicroscoop. (a) Overzichtsbeeld: (D) dorsale zijde; (V) begin van de ventrale groeve. Schaalbalkje = 100µm. (b) Detailbeeld: (1) labrum, (2) mandibel, (3) maxilla, (4) hypopharynx, (5) maxillaire palp en (6) labium. Schaalbalkje = 50µm.
Met behulp van een elektronenmikroscoop ziet men dat het basispatroon van de cuticula een hexagonale schikking is van driehoekige granules met lineaire verbindingen ertussen (Massoud & Barra, 1980; Figuur 3). De granules kunnen gereduceerd zijn, zodat een relatief glad oppervlak bekomen wordt of ze kunnen samengesmolten en vergroot zijn zodat secundaire granules gevormd worden (Cassagnau & Lauga-Reyrel, 1985).
Soorten die gemakkelijk kunnen uitdrogen, hebben granules die dicht bij elkaar staan, ook de cuticula is verdikt, zodat de permeabiliteit en de transpiratie van water verminderd wordt (Cassagnau, 1993). Aanpassingen in de schikking van de granules komen blijkbaar niet voor (Eisenbeis & Meyer, 1986). Door de aanwezigheid van een epicuticulaire waslaag op de granules is de cuticula van de meeste soorten hydrofoob (Ghiradella & Radigan, 1974). De niet-bedekte holtes binnen de zeshoekige schikking zijn zo ingesloten door een waslaag. Dergelijke structuur maakt gasuitwisseling mogelijk, maar gaat verdamping tegen doordat in die holtes een bepaalde vochtigheid kan behouden worden (Joosse, 1973).
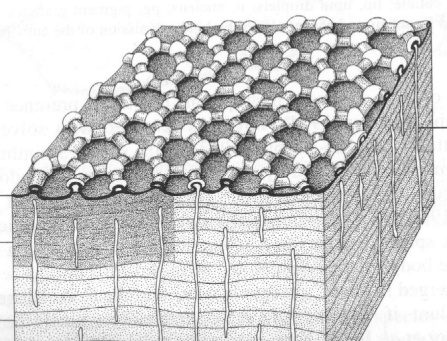
Figuur 3: Schematische voorstelling van de cuticula van Collembola. De poriekanaaltjes, die de lamellae doorkruisen, eindigen in microtuberkels (Eisenbeis & Wichard, 1987).
De meeste oppervlakte-bewonende springstaarten zijn gepigmenteerd (Noble-Nesbit, 1963a). De pigmentatiegraad is gerelateerd met de breedteligging en de hoogte, hoe dichter bij de polen of hoe hoger in de bergen, hoe donkerder de Collembola (Rappoport, 1969). Zeer gepigmenteerde soorten warmen vlugger op in de zon en worden sneller actief dan lichtgekleurde dieren (Rappoport, 1971). De pigmentatie is ongetwijfeld belangrijk als camouflage, maar vooral de bescherming tegen schadelijke UV-stralen is van belang (Zinkler & Wilking, 1989). Ook de vervuiling kan de pigmentatie beïnvloeden (Fritzlar et al., 1986).
De juvenielen lijken morfologisch heel goed op de volwassen springstaarten. Ze ontwikkelen zich paurometabolisch d.w.z. van ei tot adult door middel van een aantal vervellingen, zonder onderbreking door een larvestadium of metamorfose. Vervellingen laten in eerste instantie de groei van de dieren toe. Springstaarten vervellen gedurende gans hun leven, doch adulte collembolen veranderen noch in grootte, noch morfologisch, de vervellingen houden verband met de excretie van afbraakproducten. Tijdens de vervellingen scheiden de springstaarten een vloeistof uit, die de ruimte tussen de oude en nieuwe cuticula opvult (Barra, 1969; Krishnan, 1969), ze bevat enzymen die de binnenste laag van de oude cuticula oplossen (Noble-Nesbitt, 1963b).
2.4. Pseudocelli
Onychiuridae bezitten talrijke circulaire structuren op kop, thorax en aanhangsels, welke gekend zijn als ‘pseudocelli’ (Christian, 1989b; Fjellberg, 1973b; Hale, 1969; Hale & Smith, 1966; Pomorski, 1993; Rusek, 1984a; Weiner & Fjellberg, 1994; figuur 4). Wanneer
geïrriteerd, scheiden deze Collembola een vloeistof uit vanuit klieren gelegen onder de pseudocelli.
De klieren bestaan uit secretorische cellen, welke ingezakt liggen in de lichaamsholte en in contact staan met de haemolymfe (Rusek & Weyda, 1981). De pseudocelli zijn gemaakt van epicuticula en zijn meestal versterkt met ribben of dikkere cuticula. Levende exemplaren van Protaphorura ‘lata’ kan men er toe krijgen defensieve vloeistof te secreteren door ze te prikken met een naald. Deze naald wordt dan ‘actief’ en zal andere springstaarten en potentiële predators afschrikken, als ze in de buurt wordt gehouden (Usher & Balogun, 1966). Onychiuridae bewaard in 70% alcohol, zijn vaak bedekt door een dunne bruine rand van defensieve vloeistof, in vaste toestand gebracht na secretie van de pseudocelli. Een afwerende secretie is vrijgegeven door de pseudocellaire secretorische cellen. De secretie is waarschijnlijk door de canaliculli in de ribben van de pseudocellaire plaat naar de mediale groeve aan het oppervlak van de pseudocelli. Wanneer aangevallen door een predator is de reactie van de dieren vrij verschillend. In alle Onychiuridae lokt zo een aanval een volvation uit, als gevolg van een toename in de bloeddruk splitst de pseudocellaire plaat en een druppel vloeistof ontsnapt door de pseudocellus naar het lichaamoppervlak. Deze druppels bevatten slechts enkele intacte haemocyten. Tijdens de passage door de pseudocellaire ring en de spleet in de plaat worden de meeste haemocyten ontbonden in kleine korrels, organellen en andere plasmatische componenten. De significante componenten in verband met de afwerende functie van de druppels zijn de vacuolen van de pseudocellaire secretorische cellen. De verdedigende reactie van de collembolen wordt gestimuleerd door volvation en vrijkomen van een grote hoeveelheid van secretie aan het lichaamsoppervlak (Rusek & Weyda, 1981).
Er zijn twee basis types van pseudocelli (Rusek & Weyda, 1981), afhankelijk van de morfologie van het oppervlak. Het ene en eenvoudige type is karakteristiek voor de subfamilie Tullbergiinae. Dit type van pseudocellus is enkel zwak gescheiden van het omringende integument. De pseudocellaire plaat is onduidelijk gedifferentieerd in één of een paar kleppen, tangentiaal of in een rozet gelegen. Het draagt dezelfde granulatie als het omliggende integument of heeft enkel primaire granulatie. Het tweede basistype van pseudocelli is kenmerkend voor de subfamilies Onychiurinae en Tetrodontophorinae. Dit type is duidelijk gescheiden door een circulaire groef met primaire en lage secundaire granules. De pseudocellaire plaat in dit type is glad, zonder granules, en draagt 6+6 tot 20+20 ribben, welke de ondiepe,mediale groeve raken.
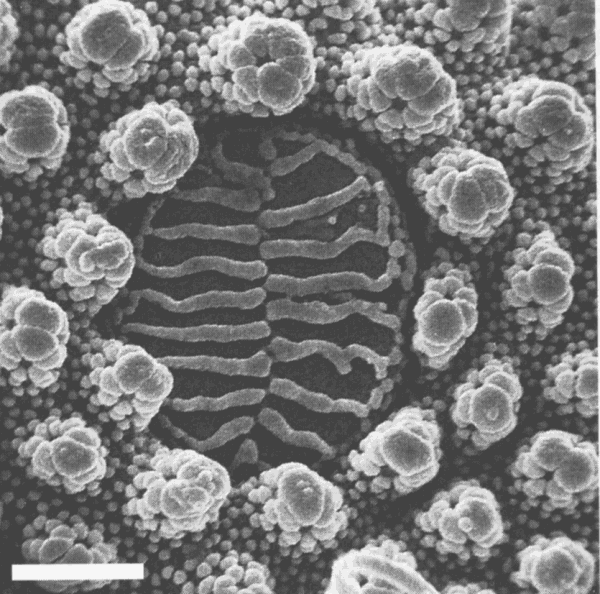
Figuur 4: SEM van een peudocellus van Protophorura sp. (Onychiuridae). Defensieve vloeistof wordt uit deze poriën geperst wanneer het dier geïrriteerd is. Het aantal en de positie van pseudocelli wordt extensief gebruikt in de taxonomie der Onychiuridae. Schaalbalkje = 3µm. (Hopkin, 1997).
Het aantal en de verspreiding van pseudocelli is belangrijk bij determinatie van de verschillende soorten Onychiuridae (Gisin, 1960a; Pomorski, 1986, 1990c, 1993; Pomorski & Skarzynski, 1989).
Uit onderzoek blijkt dat Collembolen in het algemeen reageren op smaak, reuk, licht, aanraking en geluidstrilling. Sommige soorten hebben één of meerder van deze zintuigen verloren in de loop van hun evolutie (bv. sommige bodem- en grotbewonende soorten zijn blind). Springstaarten kunnen ook reageren op veranderingen in luchtdruk (Meier et al., 1988), in vochtigheidsgraad, in temperatuur en in zuurstof- en koolstofdioxideconcentratie.
De antennae zijn rijkelijk bezet met sensorische structuren, vooral aan de top (Slifer & Sekhon, 1978). De antennen worden in de lucht heen en weer gezwaaid en tikken voortdurend tegen de grond om respectievelijk vluchtige chemicaliën en stimuli van substraat op te vangen.
De antennale segmenten dragen normaal setae, waarmee luchtstromingen en vibraties worden waargenomen, en dunwandige chemoreceptorische setae of sensilla. De meeste springstaarten hebben op hun derde antennale segment een sensorisch complex, bestaande uit twee of meerdere sensilla, die beschermd worden door setae (Gough, 1973). De top van de antenne draagt een complex van sensorische haren en receptoren (Massoud & Delamare-
Deboutteville, 1969). Zo een complex is vaak beter ontwikkeld bij soorten die in grotten leven (Figuur 5a).
Bij de meeste springstaarten komt net achter de basis van iedere antenne een postantennaal orgaan (Figuur 5b) voor. De vorm kan uitwendig verschillen van soort tot soort, maar de inwendige structuur vertoont overeenkomsten tussen de verschillende families. De functie van het postantennaal orgaan is niet goed gekend, maar het zou gevoelig zijn voor geur, vochtigheidsgraad of temperatuur (Altner & Thies, 1976).
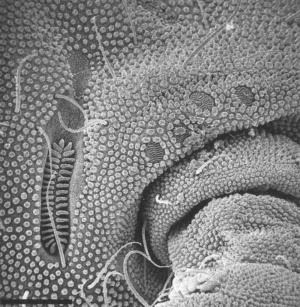

Figuur 5: (a) Elektronenmicroscoopopname van de antenne van Protaphorura sp. (Onychiuridae). Het sensorische deel is duidelijk zichtbaar op het distaal deel van de antenne. (b) Elektronenmicroscoopopname van de regio aan de antennale basis van Protaphorura sp. (Onychiuridae). De lang gerinde stryctuur aan de linkerzijde is het postantennaal orgaan. De vier smalle, ovale structuren zijn pseudocelli.
2.5.4. Ogen
Springstaarten hebben gewoonlijk een maximum van acht ocelli aan iedere kant van de kop, toch kan een extra ‘afwijkende’ ocellus aanwezig zijn (Christian & Schaller, 1982; Mitra, 1972). Epi-edafische collembolen vertonen zelden een reductie in het aantal (zie figuur 6), grot- en eu-edafische springstaarten daarentegen hebben gereduceerde ogen en zijn vaak blind (Barra, 1973; Thibaud, 1976a). De positie van de ogen en het aantal is een belangrijk kenmerk voor determinatie.
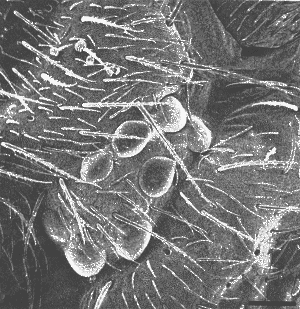
Figuur 6: SEM van het oog van Isotomurus palliceps (Isotomidae) met acht goed ontwikkelde ommatidia. Schaalbalkje = 25µm. (Hopkin, 1997).
2.6. Structuur en functie van de springvork
De springvork of furca is het meest karakteristieke kenmerkt van de Collembola (zie figuur 1). Het grotendeel van de oppervlakte-bewonende springstaarten is de furca goed ontwikkeld (zie figuren 7&8), maar bij een aantal eu-edafische soorten, die hun ganse leven in de grond vertoeven, is de springvork sterk gereduceerd of zelfs volledig verdwenen. Soorten met een grote furca hebben de neiging om één enkele, maar verre sprong uit te voeren om te ontsnappen aan predators. De hemi-edafische soorten met een gereduceerde furca maken meerdere, kleinere sprongen om dezelfde afstand te overbruggen (Bauer & Christian, 1987).
De furca is evolutionair gevormd uit een paar aanhangsels van het vierde abdominaal segment. Het aanhangselpaar is samengesmolten en vormt het manubrium, het basale deel van de springvork (Eisenbeis & Ulmer, 1987). De twee distale delen bleven apart en ontwikkelden zich tot de dentes. Het uiteinde van elke dens is voorzien van een klauwachtige structuur, de mucro (zie figuur 9). In rust houdt het tenaculum, aan de ventrale zijde van het derde abdominale segment, de furca op zijn plaats. De spieren die instaan voor het ‘springen’ zijn gespecialiseerd in uitbarstingen van anaerobe activiteit (Tosi & Ferrari, 1989; Zinkler & Schroff, 1989). Enkele sprongen na elkaar doen het ATP-gehalte in de spieren snel dalen, terwijl het ADP- en AMP- gehalte stijgt. Als de springstaart verschillende sprongen kort na elkaar uitvoert, worden de spieren zeer snel vermoeid.
 22k
22k
Figuur 7: Podura aquatica met duidlijke furca. (Driscoll)
Figuur 8: De furca, het springorgaan van Podura aqutica. (Driscoll)
Figuur 9: De mucro, de top van de furca van Podura aquatica. (Driscoll)
Collembola zijn al meer dan 400 miljoen jaar bekend op aarde en behoren tot één van de individuenrijkste dierengroepen. Springstaarten komen over gans de wereld voor in het terrestrisch milieu in vrijwel alle klimaatsgebieden en elk vegetatietype. Door hun geringe
resistentie tegen uitdroging zijn veel soorten op een leven in en/of op de bodem aangewezen. Hun verspreiding is beperkt tot de vochtige plaatsen, zodat ze bijna nooit homogeen verspreidt in hun habitat voorkomen, maar geconcentreerd op die vochtige plaatsen. Daarenboven is de vochtigheid in een bepaald habitat onderhevig aan aanzienlijke fluctuaties, zowel in ruimte, door variërende waterbindende eigenschappen van het substraat en de plaatselijke verschillen in begroeiing, als in tijd, door de heersende weeromstandigheden en de circadiane schommelingen van het dauwpunt.
De gevoeligheid voor droogteprikkels is bij de diverse soorten verschillend en afhankelijk van hun milieueisen. Vochtminnende of stenohygrische soorten vluchten al bij geringere uitdroging, dan soorten die drogere omstandigheden kunnen verdragen (Joosse & Groen, 1970). Deze gevoeligheid vertoont fluctuaties gedurende gans het leven van een individu.
Vooral tijdens en juist na de vervelling is het gevaar voor uitdroging zeer groot, omdat de nieuw gevormde cuticula nog geen verhardingsproces heeft doorgemaakt en daardoor een hoge transpiratie toestaat. Dit proces zal dus moeten plaatsvinden onder de meest gunstige omstandigheden in het belang van de overleving van het individu.
Naast de heterogeniteit van water in de bodem spelen ook andere factoren, zoals temperatuur, poriëngrootte in de bodem, pH en beschikbaarheid van zuurstofgas een belangrijke rol in de distributie van Collembola.
3.2.1. Reproductiestrategieën
Vele artikels zijn gepubliceerd over de vruchtbaarheid, de ontwikkelingsduur, de vervellingsfrequentie en de levensduur van Collembola. Eu-edafische soorten die in de stabiele omgeving van de bodem leven, vertonen de tendens parthenogenetisch te zijn en slechts enkele keren een groot aantal nakomelingen voort te brengen na een lange reproductieve periode. Epi- edafische soorten zijn seksueel reproductief en zorgen meerdere keren in hun leven voor een klein aantal nakomelingen na duidelijk afgebakende reproductieve periodes, die gesynchroniseerd zijn met het klimaat (Petersen, 1980; Van Straalen en Van Diepen, 1995).
Een aantal onderzoeken hebben aangetoond dat er bij springstaarten sprake is van een alternatie tussen een reproductief instar en een voedingsinstar. De seksueel aktieve periode bleek steeds langer te duren dan de seksueel niet aktieve periode (Joosse, Burgman en Veld, 1973; Mertens en Blanquaert, 1979).
Het reproductief gedrag van Collembola is enorm gevariëerd, gaande van soorten waarbij het mannetje spermatoforen ad random afzet in de omgeving en het vrouwtje ze zoekt, tot soorten die eerst een paringsritueel uivoeren vooraleer tot paring over te gaan . Dit laatste gebeurt bij de Symphypleona waarbij het mannetje ofwel eerst de spermatoforen afzet en het vrouwtje erover trekt ofwel het vrouwtje vastgrijpt met zijn gemodificeerde antennen en met het derde potenpaar een bolletje sperma rechtstreeks overbrengt in haar genitale opening (Blancquaert en Mertens, 1977).
3.2.2. Aggregatie
Collembola kunnen geaggregeerd voorkomen tgv. abiotische factoren in hun microhabitat of tgv. chemische communicatie (Sectie 4.5.2).
Aggregaties hebben een indirect effect op de processen van bodemvorming. Collembola behoren tot de zeer heterogene groep van de afbrekers en voeden zich vooral met bodemschimmels. Soms komen in de natuur aggregaties voor van honderden tot duizenden individuen. De bodemvormingsprocessen kunnen beïnvloed worden door de densiteit en de grootte van de aggregaties, alsook de tijd dat die aggregaties in de bodem en in de strooisellagen blijven bestaan (Leonard & Bradbury, 1984).
3.2.3. De rol van Collembola in afbraakprocessen
Het wordt algemeen aanvaard dat de bodembiocoenose (bacteriën, Protozoa, fungi en dieren) de vrijstelling van nutriënten uit decomposerend strooisel bevordert. De afbraak van organisch bodemmateriaal kan opgedeeld worden in twee processen; mineralisatie en humificatie. Springstaarten spelen een belangrijke rol in beide processen. De kleinere eu-edafische soorten leven in kleine poriën, vaak dieper in de grond, waar weinig of geen fungale groei optreedt en waar zich relatief oud organisch materiaal met lage afbraaksnelheid bevindt. Ze voeden zich met humus en minerale bodempartikels. Grotere soorten leven in grotere poriën nabij het bodemoppervlak waar er wel fungale groei is en waar relatief vers organisch materiaal met hoge afbraaksnelheid voorkomt. Het zijn vaak selectieve fungivoren die zich voeden op nutritionele hyfen. Bodembiologen nemen algemeen aan dat tijdens de afbraak van nutriëntarme bodems micro-organismen de neiging vertonen om de voedingsstoffen in hun biomassa te immobiliseren, waardoor de decompositie vertraagt. Door het afgrazen van die micro-organismen versnellen springstaarten de afbraak en stimuleren ze de groei van de hyfen (Hopkin, 1989; Warnock et al., 1982). De activiteit van de kleinere soorten kan bijdragen tot de humificatie van het organisch bodemmateriaal, terwijl de grotere soorten de mineralisatie bevorderen, nl. door stimulatie van de fungale groei. Collembola zijn dus een belangrijke schakel in de biomassatransfer van organisch materiaal naar de voedselketen.
3.2.4. Interacties tussen Collembola en andere diersoorten
Springstaarten zijn prooien voor tal van andere dieren zoals spinnen, mieren, kevers, mijten, hooiwagens en pseudoschorpioenen. Collembola hebben verschillende technieken ontwikkeld om hun predators te verschalken. Onychiura hebben bijvoorbeeld een chemisch afweersysteem, ze bezitten een klevend en toxisch haemolymfe. Als dieren van deze familie aangevallen worden beginnen ze te ‘bloeden’. Langs cuticulaire openingen - pseudocelli genaamd - wordt de afwerende haemolymfe uitgescheiden. Sommige springstaarten zouden toxisch of onsmakelijk zijn voor arthropode predators, een gevolg van chemicaliën, teruggevonden in de haemolymfe van de betreffende Collembola, die onsmakelijk of verlammend zijn (Messer et al., 1998, zie 4.5.3.). Andere soorten kunnen zeer snel lopen, maar het belangrijkste ontsnappingsmechanisme is meestal het springen. Sommige predators zijn ook aangepast om beter en gemakkelijker springstaarten te kunnen vangen. Een kleverig labrum of aangepaste antennes kunnen de kans op ontsnapping sterk reduceren.
Uitwendig parasitisme blijkt zeldzaam te zijn. Miles (1971) vond nematoden die aan de oppervlakte van Orchesella villosa hingen. Bij grotbewonende Collembola komen soms ectoparasitaire fungi voor (Christian, 1990). Inwendig parasitisme komt frequenter voor, Protozoa en ook nematoden werden in verschillende soorten aangetroffen. Men kan concluderen dat niet parasitisme, maar predators de meest significante doodsoorzaak zijn van springstaarten.
4. Chemische communicatie bij dieren
Chemische communicatie is de oudste manier van communiceren. De eerste levensvormen in de oceanen hadden reeds het vermogen om chemische substanties te detecteren en de componenten, die nodig waren voor het cellulaire metabolisme, op te nemen. Het detecteren van voedsel was en is nog steeds de primaire functie van de meeste chemoreceptorische organen. Vanaf het ogenblik dat de organismen het vermogen ontwikkelden om duidelijk onderscheid te maken tussen chemische substanties afkomstig van konsoorten en de chemische componenten van voedsel, ontstond er een primitieve, sociale organisatie en communicatiesysteem. Vroege Metazoa rekenden volledig op chemische communicatie om de vrijstelling van gameten te synchroniseren en om de bevruchting tussen gelijksoortig sperma en eieren te mediëren. Meer geëvolueerde organismen ontwikkelden in korte tijd twee soorten chemische detectiesystemen, nl. reuk en smaak.
Om een chemisch signaal over te brengen, moeten de individuele moleculen de volledige afstand tussen de zender en de ontvanger afleggen.
Er zijn drie mogelijke mechanismen:
Chemische communicatie gebeurt op meerder biologische niveaus. Chemische stoffen, uitgescheiden door endocriene klieren, die de communicatie verzekeren tussen verschillende organen van eenzelfde individu, worden hormonen genoemd. De chemische stoffen die instaan voor een uitwendige chemische communicatie, dus communicatie tussen verschillende individuen, worden semiochemicaliën genoemd. De chemicaliën die intraspecifiek werken (die dus communicatie toelaten tussen dieren van eenzelfde soort) worden feromonen genoemd.
De chemicaliën met een interspecifiek effect zijn onderverdeeld in vier functionele groepen:
Er is een grote waaier van chemische stoffen die als feromonen worden geïdentificeerd. Het zijn allemaal organische componenten met een koolstofskelet. De chemische samenstelling van feromonen is afhankelijk van het type van transmissie (via diffusie, stroming of contact) en het medium (water of lucht). Stoffen die verspreid worden via de lucht moeten vluchtig zijn (Wilson en Bossert 1963; Wilson 1970). De vluchtigheid is sterk gecorreleerd met de moleculaire grootte en het moleculair gewicht: hoe groter en zwaarder de molecule, hoe minder vluchtig. De meeste vluchtige feromonen bezitten een koolstofskelet van vijf tot twintig koolstofatomen (Figuur 10). De meerderheid van die feromonen heeft een functionele groep met één zuurstofatoom, zoals een alcohol, een aldehyde of een keton. Andere feromonen zijn carbonzuren of esters met twee zuurstofatomen, ook alkanen of alkenen zonder zuurstofatomen komen voor. De verschillen bij feromonen vallen op door de plaats van de functionele groep, de aanwezigheid en positie van dubbele of driedubbele bindingen, het voorkomen van ringstructuren en door de aanwezigheid van andere atomen in de molecule zoals stikstof en zwavel.
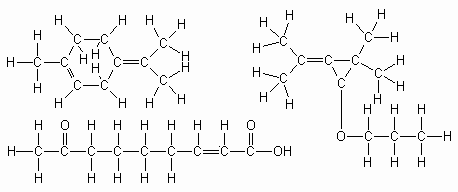
Figuur 10: Enkele voorbeelden van vluchtige, chemische geurstoffen. A: Algemeen voorkomend alarmferomoon bij termieten. B: Een seksferomoon bij een kakkerlaksoort (Periplatena americana). C: Een substantie van de honingbijenkoningin (Apis mellifera).
De restrictie op de grootte geldt niet voor contactferomonen en voor de watergetransporteerde feromonen. Organische componenten die hoofdzakelijk koolstof (MW = 12) en waterstof (MW = 1) in hun keten hebben, kunnen een kleinere dichtheid hebben dan een uit zuurstof (MW = 16) en waterstof bestaande keten, vandaar dat ze blijven drijven ongeacht hun grootte. Daarom kunnen grote organische componenten zoals lipiden en proteïnen in deze omstandigheden gebruikt worden. De moleculen die door water verspreid worden, moeten wateroplosbaar zijn en worden gedetecteerd door olfactorische receptororganen. Contactferomonen in een terrestrische omgeving kennen evenmin een restrictie op hun grootte en daardoor is er een grote variëteit aan chemische stoffen beschikbaar voor de verspreiders (Carr 1988).
4.2.2.Productieplaatsen
Feromonen kunnen afkomstig zijn uit twee verschillende bronnen:
De actieve chemische componenten zijn in het eerste geval veel gemakkelijker te identificeren. Zowel vertebraten als invertebraten bevatten talrijke klieren die bestaan uit secretorische cellen en specifieke chemische stoffen produceren. De exocriene klieren staan in voor de productie van feromonen en allomonen. Zij kunnen onderverdeeld worden in twee types op basis van de structuur en manier van secreteren:
Lichaamsopeningen, geassocieerd met de vertering en de reproductie, zijn opvallende uitscheidingsplaatsen voor chemische stoffen. Afvalproducten, zoals amines en de bijproducten van steroïdhormonen die geëlimineerd worden via de urine en de faeces, kunnen belangrijke informatie doorgeven aan andere dieren die ze detecteren. Potentieel belangrijke, vluchtige bestanddelen worden aan het spijsverteringskanaal gegeven door de lever via de gal. Exocriene klieren, die in direct contact staan met het spijsverteringsstelsel en waarvan de primaire functie het uitscheiden van verteringssappen is, kunnen secundair belangrijke feromonen produceren bv. het speeksel van mannelijke beren en egels is een bron van feromonen die gebruikt worden bij het paren (Signoret 1970; Perry et al. 1980). Ook het urogenitaalstelsel kan chemische informatie doorgeven aan konsoorten over inwendige metabolische processen.
4.3. Transmissie van afgezette signaalstoffen
Als de actieve chemische component een groot moleculair gewicht heeft en niet vluchtig is, kan het enkel gedetecteerd worden bij contact en wordt de actieve ruimte gereduceerd tot nul. De chemische samenstelling van het secreet, de eigenschappen van het afzetoppervlak en de omgevingsvariabelen hebben daardoor een grote invloed op het zendvermogen en de levensduur van het signaal.
Afgezette signaalstoffen zijn meestal vetklierproducten, waarin het actieve feromoon ingebed is in een matrix van andere componenten. De niet-actieve componenten van deze secreties zijn alles behalve inert en spelen een belangrijke rol in de regulatie van de emissie. De meer vluchtige, actieve deeltjes gebruiken het olieachtig, lipiderijk, sebum en squaleen van de klieren als dragers. De emissie van deze componenten verloopt trager naarmate de dragers meer polair zijn. De functionele groep van het feromoon beïnvloedt zijn interactie met de drager door de sterkte van de waterstofbindingen te bepalen. Een niet-polaire koolstofmolecule is minder geneigd waterstofbindingen aan te gaan met de drager dan een carboxylzuur en zal daarom sneller verdampen dan een polaire koolwaterstof. De componenten van de drager beschermen de actieve component ook tegen effecten die eigen zijn aan het substraatoppervlak en verzekeren dus een signaalmissie voor een verlengde periode.
De fysische en chemische eigenschappen van het substraat, zoals oppervlaktelaag, porositeit en chemische polariteit, hebben allen een invloed op het zendvermogen en de levensduur van het signaal (Regnier and Goodwin, 1977). De oppervlaktelaag bepaalt de beschikbaarheid van bindingsplaatsen voor het feromoon. De porositeit beïnvloedt de snelheid waarmee de chemische stof wordt vrijgelaten; poreuze oppervlakken, zoals klei vertragen de vrijlating veel meer dan houten oppervlakken. De meeste natuurlijke oppervlakken zijn elektrisch geladen, omdat ze samengesteld zijn uit polaire materialen; de meeste organische feromoonmoleculen zijn ook polair, zodat ze geneigd zijn om zich vast te houden aan het natuurlijke oppervlak, waardoor de emissiesnelheid wordt gereduceerd.
Omgevingsfactoren als de vochtigheid, de temperatuur, de wind en de zon hebben ook een invloed op de levensduur van het signaal. Hoge temperaturen en wind verhogen de verdamping en de diffusiesnelheden, waardoor de chemicaliën zich sneller verspreiden en dus sneller verdwijnen. Blootstelling aan de zon (UV), veroorzaakt chemische afbraak. Signaalstoffen zijn dus beter chemisch stabiel, zodat ze UV-straling kunnen weerstaan. De dragerscomponenten beschermen het actieve feromoon tegen degradatie, veroorzaakt door de blootstelling aan de wind en de zon. Een hoge vochtigheid en regen vormen de grootste invloeden op de duur van het signaal. Zelfs een korte blootstelling aan een gemiddelde vochtigheidsgraad kan een snelle vrijlating van het actieve feromoon veroorzaken. De reden voor deze vluchtigheid ligt bij de competitie die de sterk polaire watermoleculen aangaan voor de waterstofbinding in de drager. De verdamping van geurstoffen, die via waterstofverbindingen met de drager verbonden zijn, kan enkel geschieden wanneer deze bindingen verbroken worden. De snelle vrijlating van een signaalstof in de aanwezigheid van water stelt een goede strategie voor om de levensduur van een signaal te maximaliseren en om te verzekeren dat een feromoon enkel wordt losgelaten als er een ontvanger aanwezig is. Als de ontvanger likt, uitademt of water verliest op een gemerkt oppervlak, wordt er direct een actief feromoon vrijgelaten. De snelle degradatie van afgezette signaalstoffen door de vochtigheid van de omgeving en de regen, zal daardoor een onvermijdelijk gevolg zijn van deze zeer nuttige zenderstrategie.
4.4. De receptie van chemische signalen
Chemische receptoren kennen een eenvoudige ontwikkeling, daar chemische stimuli direct kunnen deelnemen aan biochemische reacties zonder dat er een sensorische omzetting bij te
pas moet komen. Deze directheid verklaart het universeel gebruik van chemische communicatie en zijn aanwezigheid bij de meest primitieve organismen. De meest eenvoudige en de meer geëvolueerde receptoren, of ze nu gevoelig zijn voor luchtgetransporteerde, watergetransporteerde of contactchemicaliën, hebben heel wat eigenschappen gemeen.
4.4.1. Algemene eigenschappen van chemosensorische receptoren
Hetzelfde basismechanisme wordt gebruikt om chemische stimuli te detecteren, ongeacht of de receptor nu een ééncellig organisme of een cel in een sensorisch orgaan is (Vogt en Riddiford 1986; Caprio 1988; Bruch et al. 1988). Elke stimulerende molecule bindt zich op een specifieke proteïnereceptor, die zich op het celmembraan bevindt. De receptorproteïne ondergaat hierdoor een structurele of een chemische verandering. Deze veranderingen worden dan gekoppeld aan latere biochemische processen in de cel. Vervolgens komt de stimulerende molecule los van de receptor en kan deze laatste weer binden met een ander molecule. De intensiteit van het antwoord neemt toe als de concentratie van de chemische stimulus en het aantal moleculen die gebonden zijn aan de cel stijgt. In meer geëvolueerde organen zijn de receptorproteïnen gelokaliseerd in de membraan van gespecialiseerde sensorische cellen. De binding van de stimulerende molecule, met een chemische of structurele verandering van de proteïne tot gevolg, resulteert in een depolarisatie van de cel. Dit proces kan gemedieerd worden door secundaire boodschappers in de cel. De depolarisatie heeft een invloed op de geleidbaarheid door de celmembraan en geeft het ontstaan aan een zenuwimpuls.
Chemoreceptorische cellen zijn zelf zenuwcellen of ze zijn ermee verbonden. Zoals bij andere sensorische systemen, gaat het hier om ciliaire of microvillaire cellen (Vinnikov, 1975). De cilia of de microvilli steken uit de cel en hebben zo direct contact met de externe omgeving; zij vergroten het oppervlaktegebied waar binding kan plaatsgrijpen. De aanwezigheid van ciliaire of microvillaire cellen in chemosensorische organen kent brede taxonomische patronen: de olfactorische organen van vertebraten bevatten hoofdzakelijk ciliaire cellen, terwijl de tast- en andere contactreceptoren microvillaire cellen hebben. Bij insecten zijn zowel de olfactorische als de tastreceptoren ciliair (Moran, 1987).
Chemoreceptorische cellen zijn een kort leven beschoren en worden constant vervangen. Het leven van zo ’n cel reikt enkele dagen tot enkele weken. Het systeem van nieuwe cellen die oude cellen vervangen, komt niet voor bij andere types van zenuwcellen in het lichaam. Een mogelijke verklaring voor deze vervanging is het verslijten van de chemoreceptorische membranen (Laverack, 1988). Aangezien chemoreceptieve cellen de enige sensorische receptoren zijn die met de externe omgeving in contact staan, is het niet verwonderlijk dat de gevoelige receptormembraan na een tijd beschadigd wordt van het ‘chemisch geruis’.
4.4.2. De chemosensorische systemen van Arthropoda
Bij de meeste Arthropoda kan men duidelijk twee chemosensorische organen onderscheiden: de antennen en de poot- of mondaanhangsels. De smaakreceptoren zijn gelokaliseerd aan het uiteinde van de poot- of mondaanhangsels, want deze manipuleren het voedsel. Zij hebben weinig of niets te maken met sociale communicatie.
De gepaarde antennen zijn vaak zweepachtige flagellen, die vooraan de kop uitsteken, maar sommige soorten bevatten zeer complexe antennen (Figuur 11). Mannetjes hebben soms grotere antennes dan de vrouwtjes. De antennen zijn primaire, olfactorische sensoren voor lucht- of watergetransporteerde geurstoffen, maar ze bevatten ook additionele sensoren voor smaak en tast, lucht- of waterstromingen, geluid en temperatuur. De meeste antennen bevatten tevens duidelijk gestructureerde haren, kuiltjes of haken, die sensorische cellen bevatten van ciliaire oorsprong. De algemene term voor deze sensorische structuren is sensilla. De olfactorische sensilla lijken haren, die één of drie chemoreceptorische cellen bevatten (Figuur 12) (Schneider 1964; Kaissling 1971).
De grote vederachtige antennes van veel mannelijke motten zijn ontwikkeld om de seksuele aantrekkingsstoffen van de vrouwtjes te detecteren. Er wordt geschat dat ze 80% van de moleculen die hen passeren kunnen opvangen. Het vrouwelijke feromoon bevat tussen de één en de zes gelijkaardige componenten. De feromoonspecifieke sensilla van het mannetje bevat identificeerbare cellen, die selectief gevoelig zijn voor één of een mengsel van deze componenten. De sensilla zijn zeer gevoelig (d.i. een zeer kleine K). In sommige gevallen kan zelfs op één enkel feromoonmolecule een reactie volgen.
Bij soorten met eenvoudigere en minder dimorfische antennes vormen de contactchemoreceptoren een belangrijk onderdeel van de sensilla. De dieren betasten elkaar met de antennen bij ontmoetingen (antennatie genoemd). Via receptie van niet-vluchtige componenten op het exoskelet kan er informatie verkregen worden over de soort, het geslacht en de individuele identiteit.
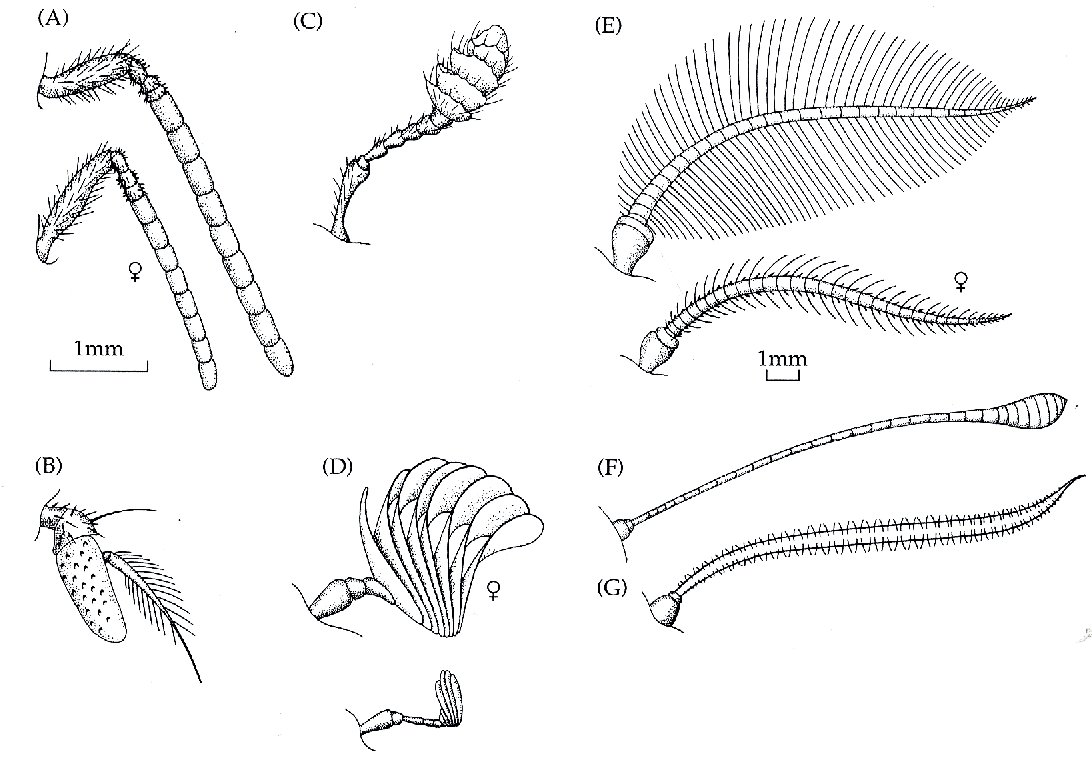
Figuur 11: Antennae van diverse insecten: (A) de mannelijke en vrouwelijke honingbij (Apis mellifera), (B) de vleesvlieg (Sarcophaga), (C) de aaskever (Necrophorus), (D) de mannelijke en vrouwelijke mestkever (Rhopaea), (E) de mannelijke en vrouwelijke nachtpauwoog (Antheraea), (F) de nummervlinder (Vanessa) en (G) de pijlstaartvlinder (Pergesa). Dezelfde schaal werd gebruikt voor A-D en E-G (naar Kaissling, 1971).
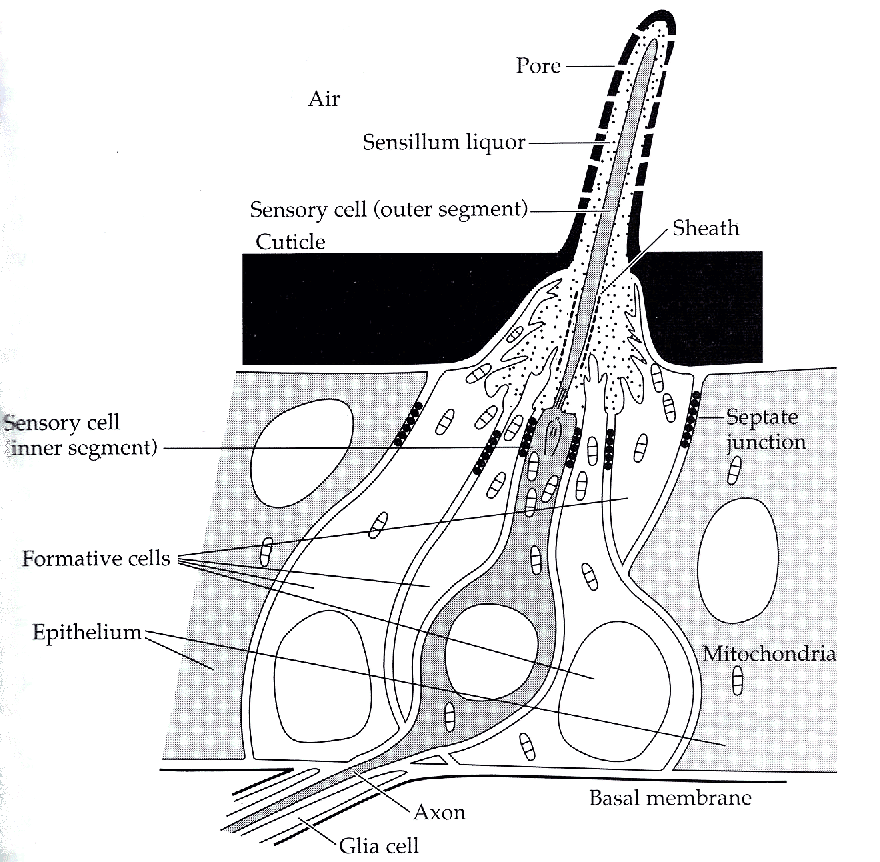
Figuur 12: Een olfactorisch sensillum van een insect. Een olfactorisch sensillum gelijkt op een eenvoudige haar, maar bij een haar zijn er poriën aan de oppervlakte, die toelaten dat olfactorische moleculen naar binnen kunnen. Aan de binnenkant baadt het uitstekend segment van één van de drie sensorische cellen in een vloeibaar medium. De sensorische cellen zijn direct verbonden met de hersenen (naar Kaissling, 1971).
4.5. Feromonen
4.5.1. Algemeen
De term ‘feromoon’ werd het eerst voorgesteld en gedefinieerd door Karlson en Lüscher (1959) als "een groep van biologisch actieve substanties, lijkend op hormonen in bepaalde
aspecten, doch niet tot hen behorend, want niet van endocriene klieren afkomstig". Deze definitie is volgens Bradbury en Vehrenkamp (1998) nog steeds geldig: "Feromonen zijn substanties, die gesecreteerd worden door een individu en gedetecteerd worden door een tweede individu van dezelfde soort. Bij dit laatste individu treedt dan een specifieke reactie op, bijvoorbeeld een bepaald gedrag of ontwikkelingsproces." Het woord zelf, afkomstig van het Griekse ‘pherein’ (= overbrengen) en ‘hormon’ (= stimuleren), werd vroeger veel bekritiseerd op etymologische gronden, maar wordt nu wereldwijd aanvaard en gebruikt in de wetenschappelijke literatuur.
Feromonen worden door een grote variëteit aan soorten gebruikt, gaande van primitieve protozoa tot de hogere primaten, als voornaamste middel om informatie over te brengen. Het wordt gesecreteerd als een vloeistof en getransporteerd als een vloeisof of gas.
Naargelang de soort en de situatie waarin een dier zich bevindt op een bepaald moment, worden bepaalde feromonen gebruikt:
Aggregatieferomonen: voor het verzamelen van mannelijke en vrouwelijke dieren om zich klaar te maken om te migreren of om de ontdekking van geschikt voedsel te melden. Om een rustige, veilige plek aan te tonen, om beide seksen samen te brengen en zo de reproductiekansen te verhogen of om de gastheerresistentie te breken door massaal aan te vallen;
Seksferomonen: trekken het andere geslacht aan;
Alarmferomonen: zorgen ervoor dat anderen wegblijven als er gevaar dreigt;
Ontmoedigingsferomonen: worden afgezet door vrouwelijke dieren, nadat ze hun eieren gelegd hebben, om andere vrouwtjes te ontmoedigen om zelf nog eieren te leggen, zodat er geen overpopulatie mogelijk is;
Territoriumferomonen: bakenen een territorium van een dier af.
Er zijn nog tal van feromonen, die gebruikt worden voor een variëteit aan andere gedragsfuncties.
Een feromoon kan direct ageren op het centraal zenuwstelsel van de ontvanger en onmiddellijk resulteren in een gedragsverandering; dit noemt men een ‘releaser’ effect. Of, een feromoon kan een fysiologische verandering initiëren of de ontvanger kan een nieuw gedragspatroon verwerven; zo ‘n feromoon geeft een ‘primer’ effect. Voorbeelden van releaser feromonen zijn die, die betrekking hebben op seksuele aantrekking, alarm en agressie, spoorproductie, aggregatie en wederzijdse herkenning. Primer feromonen gaan onder andere de inhibitie van de reproductie aan, ze spelen een essentiële rol in de organisatie en cohesie van de meer ontwikkelde gemeenschappen (vb. bij sociale bijen).
Bepaalde pestsoorten blijken enorm afhankelijk te zijn van feromooncommunicatie om te overleven. Na jarenlang onderzoek op het gewone feromooncommunicatiegedrag bij pestsoorten, kan men nu het gedrag manipuleren in het eigen voordeel. Het voorheen adaptief gedrag, zoals de oriëntatie van een dier naar een feromoonsecreterende partner, kan men veranderen in een niet-adaptief gedrag, zoals toetreden in val geïmpregneerd met hetzelfde
feromoon. Feromonen worden al een geruime tijd gebruikt als lokaas in vallen, zodat men een overzicht krijgt van de distributie en de abundantie van de pestinsecten (Shorey H.H. en McKelvey 1977).
4.5.2. Feromonen bij Collembola
Niet enkel de zoektocht naar het meest gunstige microklimaat drijft de Collembola samen, ze produceren zelf ook aggregatieferomonen (Krool & Bauer, 1987; Leonard & Bradbury, 1984; Verhoef et al., 1977a, b). De belangrijkste taak van deze feromonen zou het verzekeren zijn van ontmoetingen tussen de volwassen dieren van eenzelfde soort. Dit is voornamelijk van belang bij zich seksueel voortplantende soorten, want spermatoforen lijken enkele uren na hun afzetting, hun vruchtbaarheid te verliezen. Het vermoeden dat enkel de volwassen dieren aggregatieferomonen produceren, wordt bevestigd door Krool & Bauer (1987). Die aggregaties, die dus een beschermende functie met betrekking tot overleven hebben, vergroten dus ook de reproductiekansen.
De aggregatieferomonen zijn niet strikt soortspecifiek, hoewel alle reeds geteste soorten meer attractie vertonen voor hun eigen feromonen, dan voor feromonen aangemaakt door andere soorten. Een bepaald feromoon zou bij sommige soorten het gedrag synchroniseren, in die zin dat het een simultane vervelling induceert (Leinaas 1983b). Andere soorten kunnen bij gevaar een alarmferomoon uitscheiden, waardoor de activiteit van de dieren stijgt en dus waarschijnlijk ook hun kans op ontsnapping vergroot (Purrington et al. 1991). Er zijn feromonen aanwezig in de faecale proppen en het spijsverteringskanaal is waarschijnlijk de primaire secretieplaats (Verhoef, 1984). Misschien zijn ook de anale zakjes een bron van feromonen (Leinaas 1988).
4.5.3. Chemisch afschrikwekkende stoffen in poduroide Collembola
Het wordt algemeen aangenomen dat alle poduromorfe collembolen substanties bezitten die predators afschrikken en veel bioassays ondersteunen deze veronderstelling (zie tabel 1). Vier fenolische substanties werden gevonden in Neanura muscorum (zie figuur 13), welke verondersteld werden een rol te spelen als afschrikwekkend middel tegen predators. In de bioassays met Peragamasus norvegicus (Acarina, Parasitidae), een grote predatorische mijt die zich voedt met Collembolen, werden twee gedragscategorieën opgemerkt, als indicator voor de aanwezigheid van afschrikkende stoffen: eten komt niet of voor slechts een korte periode voor en het uitgebreid schoonmaken van de sensorische organen met welke de potentiële prooi getest is in de aanwezigheid van afwerende substanties. Het resultaat leidt naar de conclusie dat 2-aminofenol een afwerend effect heeft op P. norvegicus. Dit zou de hoofdrol van de substantie voor N. muscorum in het veld kunnen zijn. De substantie 2-aminofenol komt slechts in kleine hoeveelheden voor in de collembol, terwijl de meest voorkomende aromatische verbindingen, 2,4-dimethoxyaniline en 1,3-dimethoxybenzeen geen afschrikwekkend effect tegen de predatorische mijt vertonen.
Figuur 13: Aromatische substanties aanwezig in de collembol Neanura muscorum. Respectievelijk fenol, 2-aminofenol, 1,3-dimethoxybenzeen en 2,4-dimethoxyaniline.
2,4-Dimethoxyaniline blijkt niet toxisch te zijn en enkel lage antimicrobiële activiteit te vertonen; tot nog toe is er geen duidelijk idee van wat de rol is. Maar aangezien het in grote hoeveelheden aanwezig is, in niet enkel de springstaarten maar ook in hun eieren, is het
onwaarschijnlijk slechts een metabolische intermediair te zijn. Het zou een rol kunnen spelen als antimicrobiële vertegenwoordiger of dienen als inactieve pool voor de generatie van 2-aminophenol. 1,3-Dimethoxybenzeen, daarentegen lijkt zeer toxisch en lijkt de juiste substantie als afweer. Dit werd nochtans niet aangetoond bij de experimenten. Een andere eigenschap van deze verbinding is zijn sterke geur, welke zelfs door de mens kan waargenomen worden van een enkele dode N. muscorum, blijkt gebruikt te worden door de collembolen. Het brengt een primitieve alarmreflex teweeg bij de springstaarten.
Het defensieve mechanisme in Podura aquatica vertoont zeer verschillende efficiënties afhankelijk van de predator waartegen het werkt. Microvelia reticulata (Heteroptera, Veliidae) valt de springstaart aan en penetreert de cuticula van Podura in de meeste gevallen, maar voedt zich duidelijk korter in vergelijking met zijn gedrag bij Folsomia candida. De kever Stenus comma, daarentegen lost Podura aquatica voordat de springstaart serieus beschadigd is. Gereduceerde voedingstijd kan geïnterpreteerd worden als zijnde een afschrikkend effect, maar het dient niet tot het dier dat aangevallen werd, daar de collembol meestal de aanval niet overleeft. Het is mogelijk dat Microvelia zich weerhoudt van verdere aanvallen op gelijkaardigen voor een bepaalde tijd, zodat predatordruk gereduceerd is zelfs zonder lang-termijngeheugen, aangezien Podura gewoonlijk in grote aggregatie voorkomt.
Andere predators van P. aquatica zijn kleine vissen, die zich voeden aan het wateroppervlak, zoals Gambusia affinis (Teleostei, Cyprinodontidae), welke gebruikt worden voor de controle van Anopheles larven in Zuid-Europa. Ook hier werd een afschrikkend effect gevonden van P. aquatica, welke slechts zelden opgeslokt werd zonder onmiddellijk terug uit te spuwen. Dit kan zelfs het leven redden van de aangevallen collembol, maar het waarschijnlijke leerproces voor G. affinis om deze prooi te vermijden, lijkt belangrijker. Het zou versterkt moeten worden door de opvallende kleur van P. aquatica.
Rana temporaria (Amphibia, Ranidae) wordt niet beïnvloed door Podura aquatica. Het lijkt dat het defentiemechanisme van Podura aquatica niet effectief is tegen amfibiën.
De sleutel van de chemische verdeging bij Podura aquatica kan liggen bij het ongewone tetraterpeen, Poduran, dat ontdekt werd in de springstaart. Deze substantie is nog nooit eerder in de natuur gevonden, zodat er geen informatie over zijn biologische effecten voorhanden is.
Tabel 2: Collembolen soorten beschreven als toxisch of onsmakelijk voor arthropode predators. Het betreft enkel studies die experimenten omvatten om de veronderstelde onsmakelijkheid te bewijzen (Messer et al., 2000).
|
Collembola |
Predator |
Reactie van predator |
|
Onychiurus armatus s.l. |
Chelifer cancroides |
Afschrikking |
|
Onychiurus armatus s.l. |
Scutigera coleoptrata, Lithobius forficatus, Obisium muscorum, Chthonius spec., Lycosa hortensis, Lycosa lugubris, Tarantula trabalis, Tarantula cuneata |
Afschrikking, prooi loslaten, schoonmaken |
|
Onychiurus armatus s.l. |
Lihobius spec. |
Prooi loslaten, Schoonmaken |
|
Onychiurus armatus s.l. |
Pergamasus misellus |
Verlamming |
|
Onychiurus armatus s.l., Onychiurus fimetarius, -waterig hemolymfe extract |
Myrmica laevinodis, Lepothorax tuberum, Tetramorium caespitum, Lasius niger, Lasius flavus, Formica fusca, Formica rufa, Camponotus ligniperda |
Afschrikking, Verlamming |
|
Onychiurus armatus s.l., Onychiurus fimetarius |
Mimici laevinodis, Lepothorax tuberum, Tetramorium caespitum, Lasius niger, Lasius flavus, Formica fusca, Camponotus ligniperda |
Afschrikking, Verlamming |
|
Onychiurus armatus s.l., Metalen naald bedekt met secretie |
Pergamasus lapponicus, Veigaia nemorensis |
Afschrikking, toenemende bewegingsactiviteit |
|
Onychiurus armatus s.l. |
Stenus bipunctatus-larva, Stenus biguttatus-larva, Imago, Stenus ater-larva, Stenus incrassatus-larva, Imago, Stenus boops-larva, Imago |
Prooi loslaten, Schoonmaken |
|
Onychiurus armatus s.l. |
Apochthonius minimus |
‘weigert te eten’ na contact |
|
Onychiurus fimetarius |
Lasius emarginatus |
Prooi loslaten, Schoonmaken |
|
Tullbergia krausbaueri |
Dendrolaelaps rectus |
Schoonmaken |
|
Tetrodontophora bielanensis |
Platyseius montanus |
Verlamming |
|
Tetrodontophora bielanensis |
Notiophilus biguttatus |
Schoonmaken, Desoriëntatie |
|
Neanura muscorum |
Strumigenys lousianae |
Geen aanval |
|
Neanura muscorum |
Neobisium muscorum |
Geen aanval |
|
Neanura muscorum |
Lasius emarginatus |
Gewoonlijk geen aanval, loslaten prooi, Schoonmaken |
|
Anurida maritima |
Lasius emarginatus |
Loslaten prooi |
|
Podura aquatica |
Stenus comma |
Loslaten prooi |
|
Podura aquatica |
Stenus incrassatus, Stenus boops |
Loslaten prooi, Schoonmaken |
|
Podura aquatica |
Microvelia retuculata, Hydrometra stagnorum, Pirata sp. |
Afschrikking |
|
Hypogastrura viatica, Hypogastrura sigillata, Hypogastrura unungiculata |
Myrmica laevinodis, Leptothorax tuberum, Tetramorium caespitum, Lasius niger, Lasius flavus, Formica fusca |
Afschrikking, Verlamming |
|
Hypogastrura viatica, Hypogastrura sigillata, Hypogastrura bengtssoni, Hypogastrura ununguiculata –waterig hemolymfe extract |
Myrmica laevinodis, Leptothorax tuberum, Tetramorium caespitum, Lasius niger, Lasius flavus, Formica fusca,Formica rufa, Camponotus ligniperda |
Afschrikking, Verlamming |
|
Pseudisotoma sensibilis, Folsomia fimetaria, Xeniella humicola, Xeniella grisella |
Stenus bipunctatus, Stenus biguttatus, Stenus ater, Stenus incrassatus, Stenus boops, larven en adulten |
Loslaten prooi |
|
Mesachorutes quadriocellatus |
Lasius emarginatus |
Afschrikking |
|
Hypogasrura denticulata |
Pergamasus longicornis |
Kenmerkende positie van de voorste poten, loslaten prooi |
|
Pseudisotoma sensibilis |
Stenus bipunctatus, Stenus biguttatus, Stenus ater, Stenus incrassatus, Stenus boops, larven en adulten |
Loslaten prooi, Voedt zich enkel, wanneer zeer hongerig |
|
Folsomia fimetaria |
Stenus bipunctatus, Stenus biguttatus, Stenus ater, larven en adulten, Stenus incrassatus, Stenus boops, adulten |
Loslaten prooi, voedt zich enkel, wanneer zeer hongerig |
IPM of geïntegreerd pestbeheer is zowat de beste manier om de groeiende wereldpopulatie te voeden, vanuit zowel een politiek als sociaal oogpunt. Tot op heden verliest men 50% van de totale landbouwproductie over gans de wereld. Petsbeheer kan de schade, berokkend door insecten, aan landbouwgewassen sterk verminderen. Maar het steeds stijgende gebruik van conventionele pesticiden leidt tot resistentie van de dieren, wijziging in het ecologisch evenwicht, milieuverontreiniging en uiteindelijk zal ook de economie hieronder lijden. Het aantal insecten dat resistent wordt tegen de insecticiden stijgt, waardoor de chemische bedrijven aldoor maar nieuwere en krachtigere formules moeten ontwikkelen.
Om problemen veroorzaakt door conventionele pesticiden op te lossen werd het IPM-concept bedacht. IPM combineert de chemische, de biologische en de agrotechnische aanpak om pesten te controleren. Beperkte kostprijs, minimale schade aan het milieu en maximaal effect zijn de doelstellingen.
De pest wordt eerst geïdentificeerd, gelokaliseerd en bepaald in welke mate ze verspreid is. Daarna wordt besloten welke methode, afhankelijk van de situatie, men zal gebruiken. Biologische en afgeleide methoden maken gebruik van gekende natuurlijke vijanden en van parasieten om de pest te vernietigen, maar laten onschadelijke insecten met rust. Feromonen worden gebruikt om insecten massaal in een val te lokken of om hen in ‘verwarring’ te brengen. Als deze methoden niet mogelijk blijken of als ze niet effectief genoeg zijn, dan worden de ‘grote kanonnen’ ingezet: de pesticiden (Tabel 3).
Feromonen zijn soortspecifiek en kunnen dus gebruikt worden om de grootte van een pestpopulatie te bepalen. Men plaatst vallen met een gepast seksferomoon in het geteisterde veld, afgeschermd van het licht. Zo verkrijgt een landbouwer betrouwbare informatie over de grootte van de pestpopulatie, waar zij zich bevindt, waar zij voor het eerst voorkwam, enz. Zo weet de landbouwer wanneer, waar, wat en in welke concentratie hij moet sproeien. Het gebruik van conventionele pesticiden wordt met deze methode niet uitgesloten, maar wel de hoeveelheid ervan.
Hier wordt er van uitgegaan dat als er genoeg individuen gevangen worden, de bevruchting en de productie van nieuwe generaties zwaar verhinderd wordt. Het gebruik van pesticiden wordt hier bijna volledig uitgesloten. In Noorwegen, eind jaren zeventig en begin jaren tachtig, had men problemen met de sparrenschorskever (Scolitidae; Ips typographus). Deze keversoort vernietigde miljoenen sparren en zorgde voor een economische ramp. Men plaatste honderdduizend vallen, die de vorm hadden van een boomstam en duizenden kevers konden vasthouden. Het gebruikte aggregatieferomoon (ontdekt in 1977) bestaat uit drie componenten en trekt beide seksen aan.
De insecten worden gelokt naar centraal geplaatste dozen of kokers, waarin insecticiden, met seks- of aggregatieferomonen. De feromonen worden verspreid met een soort sproeier. De efficiëntie van de chemicaliën wordt hierdoor verhoogd en de milieuverontreiniging geminimaliseerd.
Normaal worden individuen aangetrokken naar een feromoonbron, tegen de wind in, van een zone met lage concentratie naar een zone met hoge concentratie. Als men een kunstmatige zone creëert waarin de feromoonconcentratie constant is, uniform, en hoger dan de natuurlijke concentratie, dan kunnen de individuen de natuurlijke feromoonbronnen niet meer vinden en zullen ze zich niet kunnen voortplanten.
De hierboven besproken toepassingen zijn allemaal gebaseerd op seks- en/of aggregatieferomonen, maar ook andere feromonen kunnen nuttig zijn. Alarmferomonen kunnen gebruikt worden om de insecten weg te houden en de ontmoedigingsferomonen kunnen de productie van nieuwe generaties afremmen.
In bijlage II bevindt er zich een overzicht van aggregatieferomonen, typerend voor verschillende diersoorten, waarvan men de chemische formules achterhaald heeft. Dit overzicht is gebaseerd op publicaties vanaf het jaar 1992 tot het jaar 1999.
Kunnen insecten resistent worden tegen hun eigen feromonen? Men denkt het niet. Feromonen zijn essentieel voor de communicatie, de informatieoverdracht, de bevruchting en de voortplanting van de insecten; daarom is het bijna onmogelijk dat de dieren volledig ongevoelig worden voor die substanties.
De dieren zouden wel de feromoonbehandeling kunnen omzeilen. De behandelde pestpopulaties wordt nooit 100% uitgeroeid. De overblijvende individuen kunnen dan generaties voortbrengen die bijvoorbeeld andere feromonen gebruiken om te communiceren. Bij hoge concentratie seksferomonen in de omgeving zouden de dieren zelf hun feromoonsecretie kunnen verhogen, zodat ze toch de andere sekse kunnen aantrekken.
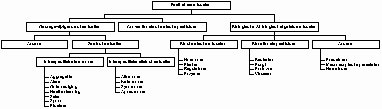 Tabel 3: Voorgestelde classificatie van pestbeheermiddelen, waarbij vooral de nadruk wordt gelegd op de biologische middelen en de chemische communicatie stoffen.
Tabel 3: Voorgestelde classificatie van pestbeheermiddelen, waarbij vooral de nadruk wordt gelegd op de biologische middelen en de chemische communicatie stoffen.
Go to the
first,
next,
last
chapter,
overview.
 20k
20k 32k
32k