2. Biologie van de collembola
2.1 Inleiding
Collembola of springstaartjes zijn de meest voorkomende Hexapoda op aarde (één liter grond bevat 200-1000 exemplaren)(Gisin,1942). Maar door hun geringe grootte (0.2-10 mm) en onopvallende levenswijze, blijven ze vrijwel ongekend. Vandaag de dag worden ze steeds minder beschouwd als primitieve vleugelloze insekten, maar eerder als Hexapoda. De Hexapoda bevatten de Collembola, Diplura, Protura en de insekten. Alle insekten zijn dus Hexapoda maar niet alle Hexapoda zijn insekten (Gordon,1999).
Het oudste fossiel dateert van zo een 400 miljoen jaar geleden (Onder-Devoon). Momenteel zijn ongeveer 7000 soorten gekend en men schat dat dit aantal kan oplopen tot 50000. John Lubbock gaf in 1871 voor het eerst de naam 'Collembola' aan deze groep. Collembola is afgeleid van ‘colle’, wat lijm betekent en ‘embolon’, wat staat voor zuiger en verwijst naar de ventrale tubus, die als vasthechtingsapparaat zou dienen op gladde oppervlakken (Maddison, 1998).
De Collembola zijn onderverdeeld in twee subordes, de ‘Arthropleona’ die min of meer langwerpig zijn, met een duidelijk onderscheid tussen thoracale en abdominale segmenten en die meestal bodembewoners zijn; en de ‘Symphypleona’ die een ronde, meer compacte lichaamsvorm hebben waarbij de thoracale en de eerste vier abdominale segmenten gefusioneerd zijn en die zelden bodembewoners zijn (fig.2.1).
Deze organismen zijn in vele terrestrische systemen vertegenwoordigd, ze bevolken onze planeet al miljoenen jaren en vormen dus één van de meest succesvolle Arthropoda.
2.2 Morfologie
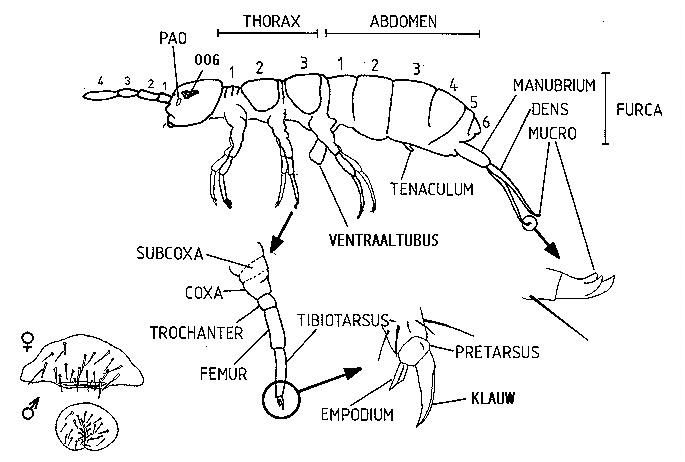
Collembola zijn kleine, vleugelloze Hexapoda. Hun lichaam bestaat uit een cephalon, een thorax en een abdomen (fig.2.2). Ze zijn bedekt met haren (setae) of schubben (gemodifieerde haren) van verschillende vorm. Vaak is het aantal en de schikking hiervan karakteristiek op soortniveau.
De kopstreek wordt gekenmerkt door interne monddelen, primair vierledige antennes, ogen en twee postantennale organen. De monddelen zijn samengesteld uit een paar inwendige mandibels en maxillen. De mondholte wordt afgesloten met het labrum. Het entognaath zijn van de monddelen van Collembola zou volgens Manton (1964) onafhankelijk zijn ontstaan van de Diplura en de Protura. Daarom hecht ze weinig fylogenetisch belang aan de groep Entognatha. Andere auteurs zoals Hennig (1969,1981) en Kristensen (1981) maken nog steeds onderscheid tussen Ectognatha (Thysanura en Pterygota) en Entognatha (Protura, Diplura en Collembola) (fig.2.3).
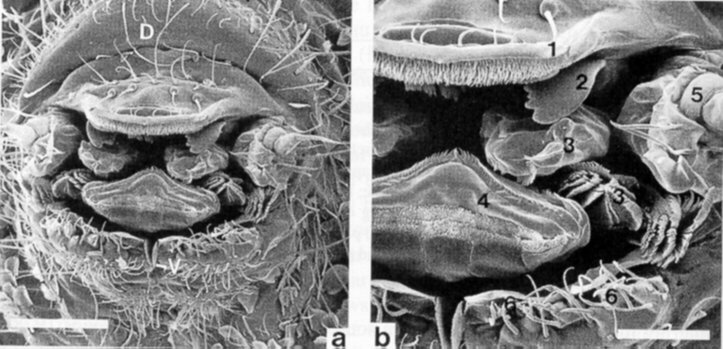
De antennale segmenten kunnen nog onderverdeeld zijn in subsegmenten. Ze zijn ingeplant op een verheven deel van de kop, met name op de antennale basis. De ogen bestaan uit sferische ocelli en een onderliggend donker pigment. Het aantal ocelli varieert van nul tot acht aan elke zijde. Tussen de ogen en de antennale basis komt vaak een sensorisch orgaan voor. Dit is het postantennaal orgaan dat bestaat uit ofwel een enkel ovoïd lichaam ofwel uit meerdere lichamen van variërende vorm en samenstelling (Fjellberg, 1980).
De thorax bestaat uit drie segmenten met telkens één paar poten. Deze bevatten slechts vier segmenten: subcoxa, coxa, femur en tibiotarsus. Het laatste segment draagt het empodium en een klauwtje. Het eerste thoracale segment kan secundair gereduceerd zijn tot een smalle, meer beweeglijke strook.
Het abdomen van springstaarten bezit slechts zes segmenten. Ze beschikken over twee gedifferentieerde abdomenaanhangsels. Het eerste, de furca, is ingeplant op het vierde abdominale segment en is een typisch spring-apparaat. Deze furca of springvork bestaat uit een basaal deel, het manubrium met daarop twee armen, de dentes, die eindigen in een klauwachtig aanhangsel, de mucro. De springvork wordt vrijwel alleen in geval van verontrusting gebruikt en maakt dan een bijzonder efficiënte vlucht mogelijk. In rust wordt zij op haar plaats gehouden door het retinaculum of hamula, een haakvormige structuur op het derde abdominale segment. De furca kan ook gereduceerd of afwezig zijn. Het tweede abdominaal aanhangsel is de ventraaltubus. Dit is een enigszins uitstulpbare, zeer dunwandige blaas die zich op de ventrale zijde van het eerste abdominale segment bevindt. Hij fungeert als zuignap bij de voortbeweging over gladde oppervlakken maar de voornaamste functies zijn de regulatie van de waterhuishouding en de ademhaling. Collembolen missen namelijk, op enkele uitzonderingen na, een ademhalingssysteem in de vorm van tracheeën. Respiratie vindt plaats door de ventrale tubus en door de huid. In verband met deze ademhalingsfunctie vertoont de huid een aantal aanpassingen. Het feit dat de huid gasuitwissling moet toelaten, betekent namelijk ook dat er verdampingsproblemen kunnen optreden.
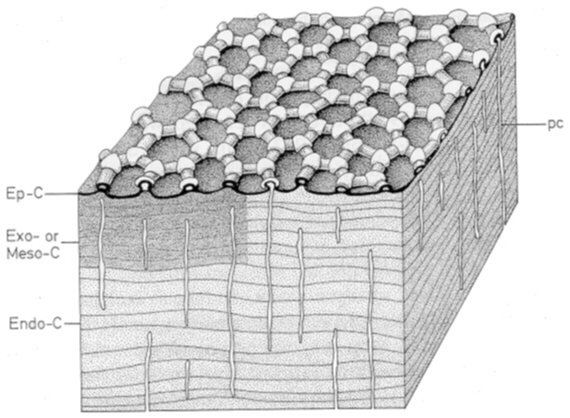
Fig. 2.4 Schematisch diagram van de cuticula van collembolen. Ep-C, epicuticula; Exo, exocuticula; Meso-C, mesocuticula; Endo-C, endocuticula. De kanaalporiën (pc) doorkruisen de lamellen van de cuticula en eindigen in de microtuberkels. Door Eisenbeis en Wichard (1987).
De cuticula vertoont vaak verhevenheden die bedekt zijn met een epicuticulaire waslaag. Deze sluiten als het ware kleine holten in die niet bedekt zijn door een waslaag. Zo een structuur maakt gasuitwisseling mogelijk maar gaat verdamping tegen doordat in de holten een enigszins vochtig microklimaat kan behouden worden (Joosse, 1973)(fig.2.4).
Morfologisch lijken de juvenielen zeer goed op de volwassen organismen. Ze maken geen larvaal stadium of metamorfose door. Opmerkelijk is dat ze continu vervellen, ongeveer één keer per week bij kamertemperatuur en dit hun hele leven lang. Die ecdyses vinden in eerste instantie plaats om groei mogelijk te maken, maar in de adulte levensfase neemt de groei sterk af (Joosse en Veltkamp, 1970) terwijl het vervellen doorgaat. Gedurende deze adulte vervellingen zullen noch de grootte, noch de morfologische karakteristieken veranderen (Thibaud, 1976). Uit verschillende studies is gebleken dat deze vervellingen voornamelijk verband houden met de excretie (Joosse, 1973). Collembolen bezitten namelijk geen buizen van Malpighi en veel van de afbraakstoffen worden in de darmwand opgeslagen. Elke vervelling wordt voorafgegaan door een voorbereidingsperiode van één tot twee dagen. Gedurende deze periode wordt niet enkel een nieuwe cuticula aangelegd, maar vindt ook een totale regeneratie plaats van het darmepitheel. Tijdens deze periode zijn de dieren inactief (Christian, 1988) en nemen ze geen voedsel op. Bij de vervelling wordt dan niet enkel de cuticula, maar ook het darmepitheel met de daarin opgestapelde afvalstoffen afgeworpen. De periode tussen twee ecdyses wordt een instar genoemd. In de adulte fase kan men een onderscheid maken tussen reproduktieve (met afzetten van spermatoforen of eieren) en niet-reproduktieve instars (voedingsinstars) (Joosse, 1981). Bij de meeste springstaartsoorten bestaat er een alternatie van reproduktieve en niet-reproduktieve instars.
Op een ecologisch verantwoorde manier kan men de collembolen onderverdelen in drie groepen. Deze indeling, zoals door Gisin (1943) werd voorgesteld, berust op een indeling naar eenvoudige morfologische kenmerken. De euedafische organismen leven in diepere bodemlagen (euedafon). Ze zijn heel klein en bezitten korte extremiteiten en antennes. Daarbij zijn ze vaak blind en pigmentloos (wit). De hemiëdafische soorten leven in de oppervlakkige strooisellaag van de bodem (hemiëdafon) en hebben tamelijk lange antennes en een goed ontwikkelde pigmentatie. Ze kunnen ook voorkomen in de bovenste bodemlaag. Epigeïsche soorten zijn voornamelijk actief in de vegetatie en op het bodemoppervlak(epigeon). Men noemt ze soms ook atmobiotische soorten (Massoud,1971). Zij hebben grotere afmetingen, een lange springvork en antennes, gevarieerde kleurtekeningen en goed ontwikkelde ocelli.
2.3 Ecologie
Collembolen komen over de gehele terrestrische wereld voor in vrijwel alle klimaatsgebieden en in elk vegetatietype; van de woestijnen tot de tropische regenwouden en zelfs in het ijs van Antarctica. Men kan ze vinden in de bodem, in bladstrooisel, mos, mest, op de vegetatie, op boomstammen, in grotten, langs de kustlijn, aan het oppervlak van poeltjes en stromen,etc. Maar de meeste soorten zijn, in verband met hun geringe resistentie tegen uitdroging, op een leven in en op de bodem aangewezen. Doordat hun verspreiding beperkt is tot vochtige plaatsen, zullen ze bijna nooit homogeen verspreid voorkomen in hun habitat. Daarbij komt dat de lokale vochtverschillen in een habitat aan aanzienlijke fluctuaties onderhevig zijn, zowel in de ruimte door de variërende waterbindende eigenschappen van het substraat en de plaatselijke verschillen in de vegetatie, als in de tijd door de heersende weersomstandigheden en circadiane dauwpuntfluctuaties.
De gevoeligheid voor droogteprikkels is bij diverse soorten verschillend en afhankelijk van de milieu-eisen: vochtminnende of stenohygrische soorten komen al bij geringere uitdroging tot vluchtbewegingen dan soorten die drogere omstandigheden kunnen verdragen (Joosse & Groen, 1970). Deze gevoeligheid vertoont fluctuaties gedurende het leven van een individu. Vooral tijdens en juist na de vervelling is het gevaar voor uitdroging bijzonder groot doordat de nieuw gevormde cuticula nog geen hardingsproces heeft doorgemaakt en daardoor een hoge transpiratie toelaat. Gedurende deze periode is het voor het overleven van de individuen dan ook vanzeer groot belang dat dit proces onder gunstige omstandigheden plaatsvindt. De nadruk moet echter wel gelegd worden op het feit dat naast of in combinatie met de heterogeniteit van bodemvocht, ook andere factoren, zoals de locatie van voedsel, temperatuur, poriëngrootte in de bodem, pH en CO2, de verspreiding van collembolen sterk kunnen beïnvloeden (Verhoef & van Selm, 1983; Mertens, 1975). Al deze factoren kunnen bijgevolg een bepalende rol spelen in de kans om andere individuen te ontmoeten en om tot reproduktie te komen.
De biologische betekenis van deze aggregaten wordt dan ook enerzijds omschreven als een ontmoetingsplaats tussen individuen om de reproduktiekansen te verhogen. Anderzijds hebben de aggregaten ook een beschermende functie met betrekking tot de overleving en de vervelling (Joosse, 1970). Tal van studies tonen namelijk aan dat bij verschillende groepen van Collembola de dieren gesynchroniseerd vervellen in de aggregaten (Joosse, 1970). Daarenboven wijzen verschillende auteurs erop dat er aanduidingen zijn dat het tot standkomen van de aggregaten veroorzaakt wordt door de produktie van feromonen (Verhoef et al., 1977; Mertens et al., 1979). Dit mechanisme van gesynchroniseerd vervellen en voorplanten is voornamelijk belangrijk voor stenohygrische soorten, het is niet noodzakelijk het algemene patroon bij Collembola. Atmobiotische soorten, die meer resistent zijn aan uitdroging, kunnen eventueel een ander gedrag vertonen dan die soorten die voor hun overleving en voortplanting aangewezen zijn op vochtigere plaatsen (Krool & Bauer, 1987).
Het optreden van aggregaties heeft ook een indirect effect op de processen van de bodemvorming. Collembolen behoren namelijk tot de zeer heterogene groep van de afbrekers (decomposers). Zij voeden zich vooral met bodemschimmels en bacteriën. Soorten die in bomen en aan de oppervlakte voorkomen eten ook algen en rottend plantenmateriaal; sommigen zijn predator. Collembolen behoren tot de weinige organismen die het insekticide DDT (dichlorodiphenyltrichloorethaan) kunnen afbreken in de bodem. In natuurlijke omstandigheden worden soms aggregaties waargenomen die uit honderden tot duizenden dieren kunnen bestaan. Daardoor kunnen de bodemvormingsprocessen beïnvloed worden door de densiteit en de grootte van de aggregaties, alsmede door de tijd dat deze in de bodem en in de strooisellaag blijven bestaan (Leonard & Bradbury, 1984).
Algemeen wordt aangenomen dat de bodemfauna de vrijstelling van nutriënten uit afbrekend strooisel bevordert. De afbraak van organisch bodemmateriaal kan opgedeeld worden in twee processen, namelijk mineralisatie en humificatie. Springstaarten spelen een belangrijke rol in beide processen. De kleine euedafische soorten leven in kleine poriën en voeden zich met humus en minerale bodempartikels. Zij leven ook veelal dieper in de bodem, waar weinig of geen fungale groei optreedt en het organisch materiaal relatief oud is met lage afbraaksnelheid. De grotere soorten leven in grotere poriën nabij het bodemoppervlak waar fungale groei optreedt en waar zich relatief vers organisch materiaal met hoge afbraaksnelheid bevindt. Het zijn vaak selectieve fungivoren die zich voeden op de nutritionele hyfen. Een vrij algemeen aanvaarde denkwijze bij bodembiologen is dat gedurende de afbraak van nutriënten-arme bodems de micro-organismen neigen tot het immobiliseren van de voedingsstoffen in hun biomassa, waardoor de decompositie vertraagt. Door het afgrazen van micro-organismen versnellen springstaarten de afbraak en stimuleren ze de groei van de hyfen (Hopkin, 1989; Warnock et al., 1982). Een hypothese hiermee in verband gesteld door Van Amelsvoort et al. (1988) is dat de activiteit van de kleinere soorten kan bijdragen tot de humificatie van het organisch bodemmateriaal, terwijl de grotere soorten de mineralisatie zouden stimuleren door de groei van fungi te bevorderen door begrazing. Collembolen vormen dus een belangrijke schakel in de biomassa-transfer van organisch materiaal naar de voedselketen (Seifert et al., 1979).
Springstaarten zelf dienen als voedselbron voor tal van andere dieren. Tot de predatoren behoren onder meer vogels, spinnen, kevers en mijten. Collembolen hebben dan ook verschillende methoden ontwikkeld om aan die predatoren te ontsnappen. Onychiuridae bijvoorbeeld hebben een soort van chemische wapens ontwikkeld onder de vorm van een klevende en toxische haemolymfe. Andere soorten kunnen zeer snel lopen. Maar bij de meeste blijkt het belangrijkste ontsnappingsmechanisme toch de mogelijkheid tot springen. Als collembolen mechanisch gestimuleerd worden, schieten ze door de actie van de furca binnen de 10-15 ms in de lucht. Dit mechanisme is enkel effectief bij soorten die op het bodemoppervlak leven. Bij de organismen die dieper in de bodem leven, is het springen als vluchtmethode vrij zinloos. Bij deze dieren neemt men dan ook vaak waar dat de furca in mindere of meerdere mate gereduceerd is. Een hypothese, door Bauer & Christian (1987) gesteld en aangetoond, is de volgende: epigeïsche soorten die voornamelijk op planten en op het bodemoppervlak leven, vertonen een vluchtstrategie die hun toelaat het bereik van een aanvaller te verlaten door één enkele, verre sprong tegen hoge snelheid. Hemiëdaphische soorten daarentegen die, afhankelijk van de vochtigheid, alterneren tussen het bodemoppervlak en de strooisellaag, voeren eerder korte sprongen uit.
2.4 Reproduktiebiologie
Collembolen zijn van gescheiden geslacht. In de meeste gevallen laat enkel de genitaalopening, gesitueerd op de ventrale zijde van het vijfde abdominale segment, het onderscheid tussen mannelijk en vrouwelijk toe. De mannelijke genitaalopening kan omschreven worden als een cirkelvormige plaat, versierd met haren en met een longitudinale spleet. Bij de vrouwtjes is de, eveneens met haren bezette, plaat eerder ellipsvormig en vertoont een transversaal gerichte spleet (Massoud, 1971)(fig.2.5).
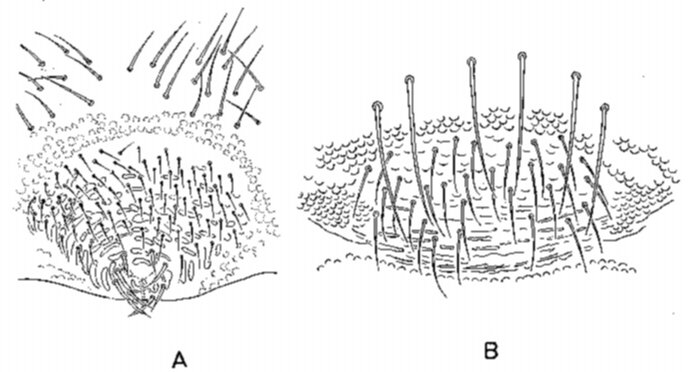
Fig. 2.5 Anurida maritima. A: mannelijke genitale plaat, B: vrouwelijke genitale plaat.
Deze primaire geslachtskenmerken zijn bij alle collembolen aanwezig, terwijl secundaire geslachtskenmerken eerder een uitzondering vormen op de regel. Bij de Sminthuridinae (Sminthurides, Sphaeridia, etc.) bijvoorbeeld, is het mannetje altijd veel kleiner dan het wijfje. Hij bezit omgevormde antennes, waarmee hij zich aan deze van het vrouwtje vasthaakt (fig.2.6).
Mannelijke individuen van het genus Bourletiellini bezitten veranderingen op de twee laatste abdominale segmenten en bij het genus Bovicornia zijn bij het mannetje omvormingen van kop en antennes te zien (Massoud, 1971)(fig.2.7).
2.4.1 Verschillende voortplantingsmethodes
Tot op heden zijn enkele gevallen van strikt parthenogenetische voortplanting bij collembolen gekend. In het algemeen zijn beide sekses bij de voortplanting betrokken, zonder dat een koppel gevormd wordt. Het mannetje zet spermatoforen af op het substraat en de vrouwtjes nemen die op.
2.4.1.1 Parthenogenese
Al sinds 1922 zette het voorkomen van populaties die enkel uit vrouwelijke individuen bestonden, enkele auteurs aan tot nadenken over de mogelijkheid van een ongeslachtelijke voortplanting bij Collembola (Massoud, 1971). Het bestaan van facultatieve parthenogenese bij collembolen werd voor het eerst in 1958 aangetoond door Choudhuri bij Onychiurus hortensis (Gisin, 1949). Sindsdien hebben verschillende studies zich over dit probleem ontfermd en men heeft het voorkomen van dit fenomeen bij verscheidene groepen van springstaarten kunnen aantonen en voornamelijk bij Folsomia candida (Willem, 1902), Folsomia cavicola (Cassagnau & Delamare, 1955) en Isotoma notabilis (Schäffer, 1896). Parthenogenese komt overwegend voor bij euedafische soorten (Berbiers, 1987). Dit is begrijpelijk, daar het leven in de bodemcapilairen geen efficiënte werking van feromonen toelaat en de visuele stimuli verhindert.
2.4.1.2 Spermatofooroverdracht
In 1952 werd door Schaller voor het eerst het bestaan van spermatoforen waargenomen (Massoud, 1971). Vandaag is hun aanwezigheid bijna bij alle soorten springstaarten gesignaliseerd. Op de bouw van spermatoforen wordt hier niet uitvoerig ingegaan, daarvoor wordt verwezen naar oa. Schaller (1970) en Betsch-Pinot (1974). Voor de manier van afzetten wordt verwezen naar Schaller (1952). Het volstaat te vermelden dat een spermatofoor bij de meeste springstaarten bestaat uit een fijn steeltje of pedunculus dat een spermadruppeltje draagt (fig.2.8). Het spermadruppeltje wordt enkel door een condensatiemembraan tegen uitdrogen beschermd. De afmetingen van zowel het steeltje als het druppeltje zijn zeer variabel (Massoud, 1971).

Fig. 2.8 Spermatofoor van Allacma fusca
Van Straalen et al. (1985) onderscheiden vijf types voortplantingsgedrag. De meeste soorten behoren tot het eerste type. De mannetjes zetten ongericht spermatoforen af, onafhankelijk van de aanwezigheid van vrouwtjes. De vorming van aggregaten door middel van feromoonproduktie (Mertens et al., 1979; Verhoef et al., 1979; Leonard & Bradbury, 1984) en het gesynchroniseerd voorkomen van de reproduktieve instars (Joosse, 1981), verhogen de kans dat een spermatofoor door een wijfje gevonden wordt. Het is niet uitgesloten dat competitieve mannetjes elkaars spermatofoor opeten vooraleer hun eigen spermatofoor wordt afgezet. Bij soorten van het epigeon is de werking van feromonen sterker dan bij soorten van het hemiëdafon (Verhoef et al., 1977). Deze verhoogde efficiëntie gaat gepaard met een duidelijker wordend seksueel dimorfisme. Bij deze soorten van het tweede type is er een verhoogde afzetting van spermatoforen in aanwezigheid van vrouwtjes. Mannelijke soorten, behorende tot het derde type zetten uitsluitend spermatoforen af in de buurt van een receptief vrouwtje. Een nog verdere verfijning van het baltsgedrag bestaat in het uitvoeren van een serie specifieke gedragingen vooraleer een spermatofoor wordt afgezet. Dit is het vierde type. Het vijfde type wordt gekenmerkt door het hoogst ontwikkeld gedrag, waarbij de mannetjes gedurende geruime tijd worden gedragen door de vrouwtjes. Dit laatste type blijkt enkel bij soorten van het epigeon voor te komen. Het is dit type waarbij rechtstreekse sperma-overdracht voorkomt.
2.4.1.3 Directe sperma-overdracht
Rechtstreekse sperma-overdracht, zonder tussenkomst van het substraat, werd waargenomen bij verschillende Sminthurides soorten en beschreven door Blancquaert (intern rapport). Het paringsgedrag van Sphaeridia pumilis werd uitvoerig beschreven door Blancquaert & Mertens (1977).

Fig. 2.9 ‘Flirtend’ paar van een mannelijke (links) en vrouwelijke (rechts) Bourletiella hortensis (Sminthuridae). Het vrouwtje is bij benadering 1 mm lang. Foto door Steve Hopkin.
2.4.2 Het afleggen van de eitjes
Een vrouwtje legt ongeveer 90 tot 150 eitjes gedurende haar leven. Dit aantal varieert van soort tot soort. De eieren kunnen ofwel individueel en verspreid afgelegd worden, zoals bij Lepidocyrtus cynaeus, of in grote eipakketten, zoals bij Isotoma viridis (Berbiers, intern rapport). Het duurt zowat een maand vooraleer de eieren uitkomen bij 8°C, maar bij hogere temperaturen gaat het veel sneller; eitjes van Tomocerus plumbeus komen uit na 3 à 4 dagen bij 20°C en de meeste specimens zullen een jaar leven.
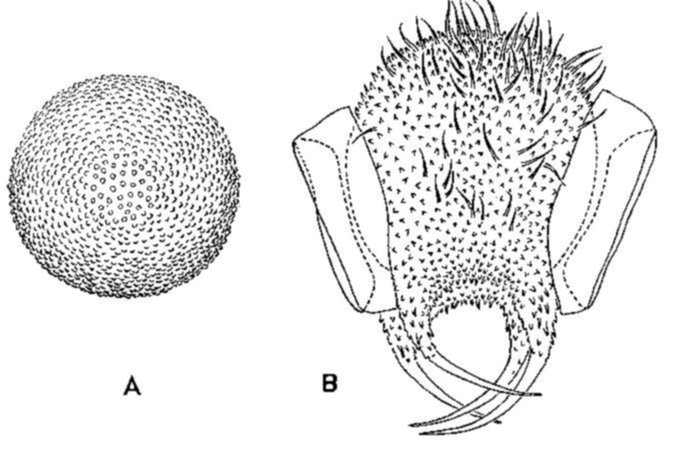
Fig. 2.10 Eitjes van Tomocerus minor. A: Sferisch ei kort na de ovipositie. B: Ei reeds voorzien van ornamenten.
Bij sommige soorten kan eikannibalisme optreden. De juvenielen ondergaan 5 tot 13 vervellingen vooraleer de seksuele rijpheid te bereiken en de tijd tussen twee ecdyses varieert met de soort en de temperatuur van 3.8 dagen in Callyntrura chibai bij 26°C tot 110 dagen bij Gulgastrura reticulosa (Gordon,1999). Sommige soorten, zoals oa. Sphaeridia pumilis, Lepidocyrtus lignorum en Isotoma viridis, leggen naast deze niet-diapauze-eieren ook diapauze-eieren af (Mertens et al.,1981; Leinaas & bleken, 1983; Berbiers, intern rapport).
Go to the
first,
next,
last
chapter,
overview.