3. Dormantie : verspreiding in de tijd
3.1 Inleiding
Elk organisme zal er steeds naar streven om zijn fitness te verhogen. Fitness is een relatieve, geen absolute term. Het aantal zaadjes van een plant of eitjes geproduceerd door een insekt, zijn geen directe maten voor hun fitness; zo dus ook het aantal nakomelingen dat ze voortbrengen niet. Het is eerder de relatieve bijdrage van een individu tot de toekomstige generaties die zijn fitness bepaalt : de fitste individuen in een populatie zijn deze die het grootste aantal nakomelingen voortbrengen relatief tot het aantal nakomelingen voortgebracht door de andere individuen van de populatie. Die individuen met het grootste aantal nakomelingen in een populatie, hebben de grootste invloed op de overerfbare eigenschappen van die populatie (Harper et al., 1996). Een organisme kan ook zijn fitness verhogen door zijn actief leven wat uit te stellen, zolang dit uitstel de kans op het voortbrengen van nakomelingen verhoogt. Dit is vaak het geval wanneer de toekomstige condities er beter uitzien dan de huidige.
Dus, een uitstel van het actief leven kan gezien worden als migratie in de tijd. In het algemeen brengen organismen deze tijd van vertraging door in een toestand van dormantie. Deze periode van sterk verminderde activiteit is voordelig omdat het organisme energie bewaart, die dan gebruikt kan worden in de periode erna. De rustfase van een organisme is dikwijls een meer tolerante toestand voor de ongunstige omgevingsfactoren; (tolerant voor droogte, extreme temperaturen, gebrek aan licht,…).
3.2 Terminologie
In dit hoofdstuk zullen enkele Engelse termen vertaald worden naar de meer gebruikelijke Nederlandse termen, ook al is deze vertaling niet officieel correct. Zo zal deze tekst geen smeltkroes zijn van Engelse en Nederlandse termen maar een éénduidig geheel vormen. Het betreft hier de termen ‘dormancy’, ‘diapause’ en ‘quiescence’, respectievelijk vertaald door dormantie, diapauze en quiescentie. Nu volgen drie auteurs die elk een verschillende terminologie hanteren.
3.2.1 Volgens Müller
De term dormantie is eigenlijk een overkoepelende term en staat voor iedere vertraging of onderbreking die optreedt in de ontwikkeling van een insekt (zowel facultatief als verplicht)(Müller,1970). Ondertussen is reeds achterhaald dat dormantie niet alleen voorkomt bij insekten, maar een fenomeen is, verspreid over het ganse planten – en dierenrijk. We kunnen ‘insekt’ in de definitie dus beter vervangen door ‘organisme’. Müller formuleerde verschillende vormen van ruststadia :
DORMANTIE
Müller,1970
Quiescentie (‘Quieszenz’) :
een facultatieve vertraging of onderbreking in de ontwikkeling als een direct antwoord, reversibel op ieder tijdstip, op het veranderen van de omgevingsfactoren van optimaal naar suboptimaal.
Diapauze :
-Oligopauze : analoog als quiescentie, maar de vertraging of onderbreking volgt echter niet onmiddellijk op de verandering van de omstandigheden, maar met vertraging. Hetzelfde geldt voor het beeïndigen van deze rustfase.
-Parapause : een noodzakelijke rusttoestand waarbij geen duidelijke inductiefase kan worden herkend. Parapause komt voor in een genetisch bepaald stadium in de ontwikkeling en is onafhankelijk van de uitwendige factoren. Voor het beëindigen ervan zijn slechts één of twee drastische veranderingen van één milieufactor nodig.
-Eudiapauze : een facultatieve rusttoestand waarbij een inductiefase kan herkend worden. Als de millieufactoren gedurende die fase niet optimaal zijn, reageert het organisme met een rusttoestand. In constante, optimale condities verloopt de ontwikkeling zonder rusttoestand. Als de rusttoestand geïnduceerd wordt door één factor, dan kan hij slechts beëindigd worden door een andere factor.
3.2.2 Volgens Harper
Harper (1977) daarentegen hanteerde een andere terminologie, meer betrekking hebbende op het onderscheid tussen plant en dier.
DORMANTIE
VOORSPELBAAR* CONSEQUENT*
DIEREN: DIAPAUZE : -facultatief -secundaire dormantie -verplicht
PLANTEN: -primaire dormantie -secundaire dormantie : =aangeboren dormantie -gedwongen -geïnduceerd
Harper (1977),*ook volgens Müller (1970)
Dormantie kan zowel voorspelbaar (‘predictive’) als consequent (‘consequential’) zijn (Müller,1970).
‘Voorspelbare dormantie’ wordt geïnitieerd nog vóór de ongunstige condities optreden en is vaak gecorreleerd met seizoengebonden veranderingen. Bij dieren spreekt men van ‘diapauze’ en bij planten wordt de term ‘aangeboren’ of ‘primaire dormantie’ gebruikt (Harper,1977).
‘Consequente dormantie’ of ‘secundaire dormantie’ daarentegen, wordt geïnitieerd als antwoord op de ongunstige omstandigheden. Dit antwoord kan zowel ‘gedwongen’ (enforced) of ‘geïnduceerd’ (induced) zijn (Harper,1977).
3.2.3 Volgens Danks
Een meer recente definiëring geeft ook het onderscheid tussen quiescentie en diapauze weer. Danks (1987) definieerde quiescentie als ‘een onmiddellijk, direct antwoord op een limiterende factor’; en diapauze als ‘een meer diepgaande onderbreking die het metabolisch programma van een organisme omleidt, weg van de directe ontwikkelingswegen en naar een duidelijkere georganiseerde pauze in de ontwikkeling (Danks, 1987).
3.3 Diapauze : voorspelbare dormantie bij dieren (naar Harper et al., 1996)
Dit ruststadium is een toestand van stopgezette ontwikkeling of groei, gepaard gaande met een sterk verminderd metabolisme en vaak gecorreleerd met seizoenale veranderingen. Dit verschijnsel is het grondigst bestudeerd bij insekten, waar voorbeelden te vinden zijn in elk ontwikkelingsstadium.
De veldsprinkhaan Chorthippus brunneus, vormt een mooi voorbeeld. Deze éénjarige soort kent een verplichte diapauze tijdens het eistadium. In deze periode zijn de eieren resistent tegen de koude winterse omstandigheden, die de nymfen en adulten ongetwijfeld zouden doden. De diapauze wordt slechts doorbroken na een lange koude periode (± 5 weken bij 0°C. of langer bij iets hogere temperatuur; Richards & Waloff,1954); dit om een vroegtijdige ontwikkeling van de eieren te voorkomen na een korte, warmere periode tijdens de winter, gevolgd door de normale, weersomstandigheden. Het brengt ook een synchronisatie met zich mee van de daaropvolgende ontwikkeling in de populatie als geheel. De sprinkhaan migreert in de tijd van de late zomer tot de volgende lente. De sprinkhaan Austoicetes cruciata daarentegen, die voorkomt in een regio van Australia met een mediterraan klimaat, legt diapauze-eieren die de droge, hete zomer overleven. Deze eieren komen opnieuw tot ontwikkeling tijdens de relatief voorspelbare vochtige winter (Andrewartha & Birch, 1954; Crawford, 1981).
De fruitvlieg Drosophila obscura, bijvoorbeeld doorloopt vier generaties per jaar in Engeland, maar slechts in één generatie treedt diapauze op (Begon, 1976). Adulten reageren op de korter wordende dagen van de herfst en betreden het diapauze-stadium. De gonadenontwikkeling van de resistente adulten wordt beeïndigd en ze zijn voorzien van grote reserves abdominaal vet. De ontwikkeling wordt voortgezet als antwoord op de langere dagen van de lente. Dus, door te vertrouwen op de volkomen voorspelbare fotoperiode, neemt Drosophila obscura een voorspelbaar diapauze-stadium aan. Dit is beperkt tot die generaties die onvermijdelijk de ongunstige omstandigheden doormaken.
3.4 Consequente dormantie bij dieren (naar Harper et al., 1996)
Er wordt verwacht dat dit type van dormantie optreedt in relatief onvoorspelbare omstandigheden. Een rustfase wordt aangenomen als direct antwoord op de ongunstige omstandigheden en wordt onmiddellijk verbroken wanneer de condities opnieuw beter zijn. Dus, de winterslaap van vele zoogdieren is een direct antwoord op de barre omstandigheden. Krachtens de energie die ze besparen bij een lagere lichaamstemperatuur, ontwikkelen ze een weerstandsvermogen en na het regelmatig ontwaken en een korte inspectie van hun omgeving, zullen ze tenslotte hun winterslaap beeïndigen wanneer de kille omstandigheden verdwenen zijn.
3.5 Zaaddormantie bij planten (naar Harper et al., 1996)
Zaaddormantie is een zeer algemeen voorkomend fenomeen bij bloeiende planten. De jonge embryo’s staken hun ontwikkeling wanneer ze nog aan de moederplant vasthangen en doorlopen een fase van verminderde activiteit. Hierbij verliezen ze gewoonlijk heel wat water en ze betreden de dormante fase in verdroogde omstandigheden. Bijna alle zaden zijn rustend als ze van de moederplant afgeworpen worden en hebben specifieke stimuli nodig om hen terug te reactiveren (ontkiemen). We kunnen drie types van dormantie onderscheiden :
1)Aangeboren dormantie (‘innate dormancy’) of primaire dormantie
is een fase waarin het embryo of het omgevende maternale weefsel absoluut een uitwendige stimulus vereist om het proces van groei en ontwikkeling te reactiveren. Deze stimulus kan de aanwezigheid zijn van water of een verandering van temperatuur, licht, fotoperiode of een geschikt evenwicht tussen licht met verschillende golflengten. Deze factoren blijken de ontkieming te synchroniseren in een bepaalde seizoensfase.
2)Gedwongen dormantie (‘enforced dormancy’)
is een antwoord op ongunstige omstandigheden. Het is een vorm van consequente dormantie. De afwezigheid van de normale vereisten voor groei (water, een geschikte temperatuur, voldoende licht en zuurstof) of de aanwezigheid van inhiberende elementen (zoals bijvoorbeeld een hoge CO2-concentratie) houden een zaad in rusttoestand. Natuurlijk bestaat de mogelijkheid dat het zaadje sterft, maar ook het tegenovergestelde is waar: zaad kan die rusttoestand voor een spectaculair lange periode behouden. Van zaden van Chenopodium album, verzameld uit archeologische opgravingen, werd aangetoond dat ze nog levensvatbaar waren na 1700 jaar (Æ dum,1965). Zaden die in rusttoestand gehouden worden, kunnen heel eenvoudig tot kiemen gebracht worden door het toevoegen van de ontbrekende bron of conditie. In de natuur kan gedwongen dormantie verbroken worden door hevige regen na een periode van droogte of door blootstelling aan de atmosfeer nabij het bodemoppervlak. Het nageslacht van één enkele plant met gedwongen dormantie kan verspreid zijn over jaren, decennia of zelfs eeuwen.
3)Geïnduceerde dormantie (‘induced dormancy’)
is een fase, tijdens een periode van gedwongen dormantie, waarin het zaad nog bijkomende eisen stelt vooraleer te ontkiemen Vele zaden van onkruid uit land-en tuinbouw zullen ontspruiten zonder enige lichtstimulus na het loslaten door de moederplant; maar na een periode van gedwongen dormantie, wordt licht een noodzakelijk element om kiemen mogelijk te maken. Deze vorm van dormantie is verantwoordelijk voor de accumulatie van enorme zaadpopulaties in de bodem. In de natuur kiemen deze zaden enkel wanneer ze aan het bodemoppervlak gebracht worden door regenwormen of door blootstelling van aarde bij het ontwortelen van een boom of wanneer er bodem omgewoeld wordt door gravende dieren of door menselijke interventie.
Een vertraging in de levenscyclus van een organisme kan een reductie van zijn reproduktievermogen betekenen en dus van zijn fitness. Anderzijds is een uitgestelde kieming vaak een ontsnapping aan dodelijke omstandigheden van de omgeving. Er moet dus ergens een compromis bestaan tussen het risico te sterven bij te vroege kieming en het verlies aan reproduktievermogen door de groei te laat te beginnen (Venable & Brown,1988).
Effectieve verspreiding in tijd is niet alleen afhankelijk van zaden met een rustfase maar ook van de levensverwachting. Zaadjes die vroeg kiemen en overleven, hebben meestal een langere groeiperiode en een hogere vruchtbaarheid dan deze die laat ontwikkelen. Bomen kunnen dan ook honderden tot zelfs duizenden jaren oud worden. Pinus aristata bijvoorbeeld, kan meer dan 2500 jaar oud worden en de datering van sommige Querqus species loopt terug tot 10000 jaar geleden. De meeste planten met zaden die lange tijd in de grond vertoeven, zijn één- of tweejarigen, waarbij de mogelijkheid tot een effectieve ruimtelijke verspreiding ontbreekt. Het betreft hier hoofdzakelijk onkruiden.
3.6 Ecologisch belang van dormantie – diapauze
Het stopzetten van de ontwikkeling (dormantie/diapauze) kan de overlevingskans van een organisme verhogen wanneer bronnen schaars zijn of de condities ver van optimaal.
De Frambozenkever, Byturus tormentosus, bijvoorbeeld is een ‘specialist’, dit wil zeggen dat zijn menu uitsluitend uit frambozen bestaat. Daar frambozen seizoengebonden zijn, zal de kever een oplossing moeten zoeken in tijden van voedselschaarste. De kever legt eitjes in de bloem en de larve vervolledigt haar levenscyclus in de zich ontwikkelende vrucht. De larve blijft dan als popstadium in diapauze totdat de framboos opnieuw bloeit, tien tot elf maanden later (Harper et al.,1996). De seizoengebonden beschikbaarheid van bronnen kan dus aanleiding geven tot het doormaken van een diapauze bij bepaalde diersoorten.
Vervolgens zijn er de niet-optimale condities, met name: verandering van abiotische factoren zoals temperatuursverhoging of – verlaging en droogte maar ook biotische zoals een grote predatiedruk, overbevolking of competitie tussen ecologisch gelijke soorten is nadelig.
3.7 Verschillende vormen van inductie en terminatie
Het is absoluut noodzakelijk dat de rustfase geïnitieerd en beëindigd wordt op het geschikte moment. Een te vroege start kan het reproduktievermogen doen dalen, evenals een te lang durende rustfase. Een vroegtijdige terminatie of laattijdige initiatie kan eveneens drastische gevolgen hebben. In alle gevallen dragen verkeerde beslissingen een hoge kost met zich mee.
Seizoengebonden diapauze wordt gewoonlijk geïnitieerd nog vóór omgevingsveranderingen plaats hebben, en dit door een cyclisch variërende aanwijzing zoals foto–of thermoperiode. Een vroegtijdige omschakeling verzekert een tijdig betreden van het diapauze-stadium voor de meeste individuen (Taylor,1980).
Onvoorspelbaar veranderende factoren, zoals vele biotische factoren, kunnen dus niet aangetoond worden met seizoengebonden veranderende stimuli, maar er wordt verwacht dat andere aanwijzingen die een hoge correlatie vertonen met de desbetreffende factor, gebruikt worden. Niettemin kunnen die gecorreleerde signalen de individuen misschien niet tijdig informeren, waardoor hun waarde daalt. Toch is er geen tekort aan voorbeelden van directe controle van dormantie.
Zo wordt "winter"diapauze bij Daphnia teweeggebracht door een verkorte fotoperiode (Stross 1971,1987) maar een plotse omschakeling met het vrijstellen van ephippia (ook wintereieren of rusteieren genoemd) tijdens de zomer wordt waarschijnlijk veroorzaakt door meer directe signalen zoals lage voedselconcentratie (Banta 1939; D’Abramo 1980), ongekende chemische factoren geproduceerd door dieren van overbevolkte populaties (Banta 1939; Carvalho & Hughes, 1983), lage temperatuur (Mortimer,1935) of andere. Lage voedselconcentratie en overbevolking zouden echter enkel aanleiding geven tot diapauze in combinatie met een korte fotoperiode (Kleiven et al.,1992). Recent werden chemische stimuli geassocieerd met vispredatie, experimenteel aangetoond. De productie van diapauze-eieren bij Daphnia magna, gestimuleerd door visexcreties, kan verklaard worden als een anti-predator verdediging die de bescherming van het genoom vrijwaart in tijden van hoge vispredatie. De chemische stimuli zijn: een chemische "alarm" stof uitgescheiden door gewonde soortgenoten en kairomonen afkomstig van de vissen zelf. Beide stoffen zijn nodig voor de productie van rusteieren, terwijl de vorming van mannelijke individuen (reproduktie controle) geïnduceerd wordt in de aanwezigheid van één of beide factoren of de afwezigheid van de factoren. De productie is natuurlijk het hoogst wanneer beide stoffen aanwezig zijn (S lusarczyk, 1999).
Fotoperiode is een zeer algemene inductie- en terminatievorm (zie 3.3) maar het is niet altijd de hoeveelheid licht die belangrijk is, ook de spectrale samenstelling kan van kapitaal belang zijn. Zo kan zaaddormantie geïnduceerd worden door licht dat een relatief grotere proportie aan straling met een golflengte van 730nm (einde rode gebied) bevat dan straling met een golflengte van 660nm (begin rode gebied). Het is precies deze bepaalde samenstelling die bekomen wordt wanneer licht gefilterd wordt door het bladerdak van een bos. Dit licht bereikt de pas afgevallen zaden die nog onvoldoende ruimte hebben om te groeien. Pas wanneer de hogere planten afsterven en het licht een andere spectrale samenstelling vertoont, daar het niet meer zo sterk gefilterd wordt, wordt de rustfase doorbroken en kan het zaad kiemen. Zo wordt dus verhinderd dat zaden ontkiemen maar geen ruimte en licht ter beschikking hebben door de aanwezigheid van overgroeiende vegetatie (Harper et al., 1996).
Vochtigheid is nog een belangrijke inductie- en terminatiefactor. Protozoa bijvoorbeeld, kunnen zich encysteren in een dormant stadium dat enkel door het terugkeren van speciale condities, zoals het heropvullen van een uitgedroogd poeltje, verbroken wordt (consequente dormantie).
Vele nematoden, voornamelijk parasieten, kennen eveneens een consequente dormantie waarbij de ontwikkeling van cysten jarenlang onderdrukt wordt en pas na een specifieke stimulus verder gaat als de omstandigheden gunstig zijn (Sunderland,1960). Deze stimulus kan de opname van de cyste door een gastheer zijn of in het geval van plantenparasieten, de vrijstelling van kiemingsstimulerende stoffen door de groeiende gastheer.
3.8 Dormantie : wijdverspreid of eerder uitzonderlijk ?
Het doormaken van één of ander ruststadium tijdens de levenscyclus is een algemeen voorkomend fenomeen in zowel het planten- als dierenrijk. Het is een onderwerp dat vaak discussies ontketend en onderzoek loopt niet altijd van een leien dakje, daar de ruststadia vaak verborgen zijn. Het is hier niet de bedoeling om alle groepen te benaderen, maar eerder een algemeen beeld te scheppen, door het geven van enkele voorbeelden.
3.8.1. Flora
Er werd reeds vermeld dat zaaddormantie een wijdverspreid fenomeen is bij bloeiende planten (zie 3.5). Bijna alle zaden zijn rustend wanneer ze door de plant losgelaten worden en leveren een significante bijdrage tot zaadbanken in de bodem. Rustende zaden komen meer voor bij éénjarigen en andere kortlevende soorten dan bij langer levende planten. Dormantie bij planten is niet beperkt tot de zaden.
Zandzegge (Carex arenia) bijvoorbeeld, maakt dormante knoppen langsheen het lineair rhizoom gedurende de groei. Deze kunnen lang nadat het rhizoom is afgestorven levensvatbaar blijven en men kan ze in grote aantallen tot 400 à 500 per m2 terugvinden (Noble et al.,1979). Prescot kervel (Chaerophyllum prescottii) heeft geen dormante zaden en geen dormante knoppen, maar dormante knollen kunnen meer dan tien jaar in de bodem overleven. Wanneer het grasland omgeploegd wordt, worden honderden rustende kervelknollen geactiveerd en aangezet tot groei (Rabotnov,1964).
Tenslotte is het gekende bladverlies van vele eeuwiglevende bomen en struiken een vorm van aangeboren dormantie. Een periode gekenmerkt door lage temperatuur en minder licht wordt doorgebracht in een bladloze, kale toestand met lage metabolische activiteit. Naast de Hogere Planten komt dormantie oa. ook voor bij algen en schimmels.
3.8.2. Fauna
Zowel binnen de ongewervelden (Invertebrata) als gewervelden (Vertebrata) zijn talloze diersoorten te vinden die een diapauze of consequente dormantie doormaken. Net zoals bij planten is ook hier dormantie niet beperkt tot het eistadium, maar de verschillende ontwikkelingsstadia en zelfs speciale structuren zoals statoblasten en gemmulae vertegenwoordigen de diapauzefase.
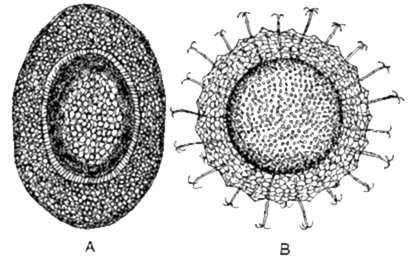
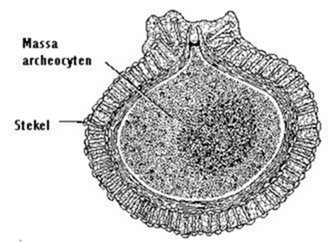
Statoblasten worden aangetroffen bij zoetwater mosdiertjes (Bryozoa). Een statoblast bestaat uit een interne massa van epidermale en peritoneale cellen, omringd door een chitineuse schaal (fig.3.1). Deze dormante organen worden in grote getale gevormd tijdens de zomer en herfst, wanneer de kolonie degenereert; ze overwinteren als resistente structuren tegen koude en uitdroging en als de lente aanbreekt kan de ontwikkeling beginnen. Een gemmula is een gelijkaardig overwinteringsorgaan: een celmassa in een omhulsel, bij zoetwater sponzen (Porifera) (fig. 3.2).
Fig. 3.1 Statoblasten van zoetwater Bryozoa.
A: Drijvende statoblast van Hyalinella punctata. B: Statoblast met haken van Crisatella mucedo.
(A, naar Rogich van Pennak. B, naar Allman)
Fig. 3.2 Doorsnede van een gemmula van een zoetwater spons. (naar Evans van Hyman).
Consequente dormantie treffen we oa. aan bij de kleinste invertebraten zoals de ééncelligen (Protozoa) en de radardiertjes (Rotifera), bij vele parasitaire vormen zoals parasitaire rondwormen (Nematoda) of weekdieren (Mollusca). De schaaldieren (Crustacea) en insekten (Insekta) zijn twee omvangrijke groepen van de geleedpotigen (Arthropoda) die momenteel veel mogelijkheden bieden qua diapauze-onderzoek. De meeste Branchiopoda (bijvoorbeeld Daphnia; Crustacea) produceren diapauze-eieren die aan intense droogte, extreme temperatuur en hoge druk kunnen weerstaan. De diapauze van vele zoetwater Copepoda (Crustacea) omvat soms rusteieren, soms larvale stadia en soms adulten. Ook de classes Malacostraca en Ostracoda bevatten enkel soorten, gekenmerkt door een rustfase in hun levenscyclus (Alekseev & Fryer, 1996). Ook vele insekten hebben te maken met een stopgezette periode van ontwikkeling en dit meestal tijdens de koude wintermaanden. Steenvliegen (Plecoptera), sprinkhanen (Orthoptera), vlinders (Lepidoptera), muggen (Diptera), wespen (Hymenoptera), kevers en torren (Coleoptera) bevatten allen enkele soorten die een diapauzestadium doormaken. Met behulp van de moderne moleculaire genetische technieken is men er in geslaagd de structuur van het diapauzehormoon bij de zijdeworm (Bombyx mori) te releveren. De diapauze in het eistadium van de zijdeworm wordt geïnduceerd door een neuropeptide
hormoon, met name het diapauzehormoon (DH). Tijdens het popstadium van dit insekt wordt dit hormoon gesecreteerd door het suboesophagiaal ganglion. De structuur van het diapauzehormoon is beschreven als een 24 aminozuurpeptide-amide waarvan de C-terminale pentapeptide-amide structuur essentieel zou zijn voor biologische activiteit (Kunio et al.,1998). Aangezien het onderwerp van deze licentiaatsverhandeling, zal diapauze bij springstaarten (Collembola; Hexapoda) uitvoerig besproken worden in het volgende puntje. Een derde groep behorende tot de geleedpotigen, de Cheliceraten, biedt nog enkele voorbeelden zoals de mijten en teken (Acarina) en de schorpioenen (Scorpiones)(Chinery, 1986).
In tegenstelling tot de invertebraten is de dormante vorm bij vertebraten voornamelijk consequente dormantie. Bij slangen (Serpentes; Reptilia) is een ruststadium waar te nemen tijdens de zomer of het droge seizoen (aestivatie). En denken we maar aan de vele zoogdieren (Mammalia) die een winterslaap houden zoals bijvoorbeeld de beer, de marmot, de egel, de vleermuis, enz.. Een opmerkelijk voorbeeld van consequente dormantie bij zoogdieren wordt ons bezorgd door die buideldieren (Marsupialia) die de ontwikkeling van een foetus verscheidene maanden kunnen onderdrukken wanneer de moeder niet veel reserves meer heeft. Zo een optie tot uitstel is niet aanwezig bij placentale zoogdieren (Placentalia)(Harper et al., 1996).
Indrukwekkender is het volgende voorbeeld, dat zowel bij buideldieren als placentale zoogdieren terug te vinden is. Embryonale "diapauze", of uitgestelde implantatie is een vorm van quiescentie, optredend in de blastocysten van zowat honderd placentale soorten en buideldieren (Renfree & Calaby,1981). De tammar wallaby (Macropus eugenii) bijvoorbeeld is in staat een blastocyst in dormantie te houden zolang er nog een buideljong gezoogd wordt. De blastocyst ondergaat geen mitose of differentiatie en kent een laag metabolisme. Hij kan opnieuw geactiveerd worden door het zuigende jong te verwijderen en ± 26 dagen later wordt de foetus geboren. Een muis is ook in staat tot een facultatieve diapauze tijdens het zogen, maar deze periode duurt niet zolang als bij de tammar wallaby (Spindler et al.,1999).
De vraag of diapauze ook bij de mens voorkomt, is voor sommigen zeer intrigerend. Maar omwille van ethische redenen kan hier echter weinig of geen onderzoek verricht worden.
3.9 Springstaarten (Collembola)
3.9.1 Adaptaties aan "abnormale" fysische condities
3.9.1.1 Diapauze
In 1964 stelde Christiansen dat nog geen echte diapauze aangetoond was bij Collembola; maar reeds in 1968 werd diapauze bij Sminthurus viridis beschreven: "Sminthurus viridis has respond to the situation by depositing special egg-batches which can not only withstand these (drying) conditions, but which must be exposed to them before the eggs can hatch the following autumn" (Wallace,1968). Walters (1968) argumenteert echter dat dit fenomeen wel als resistentie aan de droogte kan worden aanzien, maar niet als diapauze kan beschouwd worden : "This resistence of the egg to the drought is not a diapauze phenomenon but the direct induction of dormancy by unfavourable environmental conditions, as discribed by Holdaway (1927)". Ondanks deze discussie werd diapauze ondertussen experimenteel aangetoond bij de Symphypleone soorten Sminthurides aquaticus, Sphaeridia pumilis en Sminthurinus aureus (Mertens et al.,1981) en bij de Arthropleone soort Lepidocyrtus lignorum (Leinaas & Bleken,1983). Bij Anurida maritima kan diapauze afgeleid worden uit de populatiedynamiek op het terrein (Joosse,1966) en bij Tomocerus longicornis werd diapauze gesuggereerd tijdens labo-experimenten (Vegter,1987).
1. Sminthurus viridis
De rustfase bij de eieren van Sminthurus viridis kan best omschreven worden als een quiescentie: de embryonale ontwikkeling houdt op als de relatieve vochtigheid in de omgeving te laag wordt. De rusttoestand is echter onmiddellijk en op elk moment reversi
bel en de embryonale ontwikkeling herneemt wanneer de relatieve vochtigheid opnieuw toeneemt (Walters,1968).
2. Sminthurides aquaticus
De eerste dagen na het afleggen van de eieren is het onmogelijk diapauze- van niet-diapauze-eieren te onderscheiden. Maar na een 5-tal dagen (bij 20°C.) wordt een donkere vlek zichtbaar onder de eierschaal van vele diapauze-eieren (fig. 3.3). Deze vlek wordt veroorzaakt door het centraal transparant zijn van de eieren (Mertens et al.,1981). Onder experimentele condities werden van september tot juni eieren afgelegd, waarvan 80 tot 90% van het diapauze type. Onder dezelfde experimentele condities werden gedurende juli en augustus uitsluitend (100%) niet-diapauze-eieren afgelegd. Het is echter wel opvallend dat bij deze soort, evenals bij Isotoma viridis eerst niet-diapauze-eieren en nadien diapauze-eieren worden afgelegd na isolatie van op het terrein gevangen adulten. De ontwikkeling van de diapauze-eieren lijkt weinig beïnvloed te worden door de temperatuur en is waarschijnlijk onafhankelijk van de fotoperiode of de relatieve vochtigheid van het substraat (Mertens et al., 1981). Deze besluiten steunen wel slechts op één temperatuursexperiment.
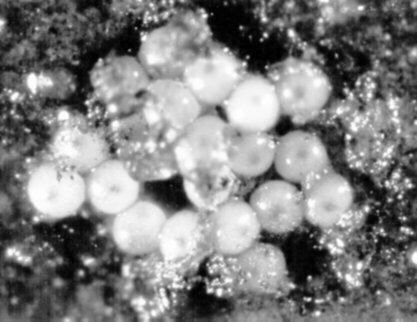
Fig. 3.3 Diapauze-eieren van Sminthuides aquaficus (door Blanquaert J.-P.)
3. Sminthurinus aureus
Ook hier worden in stockculturen eerst niet-diapauze-eieren en nadien diapauze- eieren afgelegd. Bij deze soort is echter wel een effect van de temperatuur merkbaar bij het beeïndigen van de diapauze. Mertens et al. (1981) werpen de hypothese op dat bij deze soort een eudiapauze (Müller,1970) optreedt met reductie van de fotoperiode als inducerende factor en temperatuursveranderingen als terminerende factor.
4. Sphaeridia pumilis
Bij Sphaeridia pumilis schijnt diapauze in natuurlijke omstandigheden gecorreleerd te zijn met langdurige droogte tijdens de Mediterrane zomer (Poinsot-Balaguer,1976). Onder labo-omstandigheden legt deze soort nochthans diapauze-eieren af bij een nagenoeg waterverzadigde atmosfeer. Vanaf half april worden in alle culturen onafhankelijk van de generatie in kweek, in toenemende mate diapauze-eieren afgelegd. Een endogene cyclus kan hierbij uitgesloten worden. Er rest dan nog de exogeen geregelde cyclus. Als inducerende factoren kunnen temperatuur of temperatuursveranderingen, relatieve vochtigheid, fotoperiode, voedselkwaliteit en densiteit uitgesloten worden (Blancquaert,1980). Mertens et al. (1981) besluiten dat een onbekende exogene factor de diapauze reguleert.
5. Lepidocyrtus lignorum
Ei-diapauze bij deze soort (Noorwegen) werd bestudeerd door Leinaas & Bleken (1983). Lepidocyrtus lignorum reproduceert hoofdzakelijk in de herfst. De eieren zijn van het diapauze type en de diapauze wordt beeïndigd door koude. In de lente is er een synchronisatie in het ontluiken van de eieren merkbaar, wat aanleiding geeft tot cohorte I. Enkele overwinterende adulten produceren niet-diapauze eieren in de late lente. Dit resulteert in een tweede, minder gesynchroniseerde cohorte II. Laboratorium experimenten toonden aan dat de diapauze beeïndigd wordt na een periode van lage temperatuur (2°C is reeds voldoende).
Dus, de diapauze wordt beeïndigd nog vóór het begin van de winter. Leinaas & Bleken concludeerden dat de daarna voortdurende dormantie een quiescentie is, waarbij de lage omgevingstemperatuur tijdens de winter er voor zorgt dat de eieren niet verder tot ontwikkeling komen. Het tijdstip van het uitkomen van de eieren hangt af van de stijgende bodemtemperatuur na de winter. Enkel het quiescentiestadium laat toe dat de eieren hun normale ontwikkeling voortzetten wanneer de temperatuur stijgt. De diapauze is dus niet belangrijk voor het overleven van de winter bij Lepidocyrtus lignorum. Dit fenomeen toont aan dat het belang van diapauze als overlevingsstrategie in kille, harde periodes overschat wordt. De meeste Collembola, inclusief Lepidocyrtus lignorum, ontvluchten de strenge winters door te migreren in de diepere lagen van de bodem, terwijl de meer resistente diapauze-eieren meestal naar de bovenste lagen blijven.
3.9.1.2 Adaptaties aan langdurige droogte
In streken met seizoengebonden droogte treden verschillende aanpassingenen op om de ongunstige droge periode te overbruggen. De meeste springstaarten brengen de droge zomer door in het eistadium, maar sommige soorten overleven de droge periode in het adulte stadium door een proces van dehydratatie, nl. anhydrobiose (Poinsot; 1968,1974). Gedurende dit proces worden de dieren inactief als directe reactie op de uitdroging. Ze kunnen tot 65% van hun totale watergehalte verliezen. Dit proces gaat gepaard met specifieke morfologische veranderingen in de middendarm en ultrastructurele variaties in de nucleus (Barra & Poinsot-Balanguer, 1977). Deze toestand is quasi onmiddellijk reversibel. Zodra de condities opnieuw gunstig zijn, worden de dieren weer actief (< 1uur) en hernemen in minder dan 8 dagen hun normale activiteit (Barra & Poinsot-Balanguer, 1983).
Bij een aantal andere soorten komen typische seizoenale aanpassingen voor. Deze worden gekenmerkt door zowel anatomische, morfologische, fysiologische en ethologische veranderingen. Cassagnau (1955) noemt dit syndroom ecomorfose en vooral soorten van de families Isotomidae (oa. Isotomurus palustris (Müller, 1776) en Isotoma viridis) en Hypogastruridae vertonen dit verschijnsel (Cassagnau, 1971).
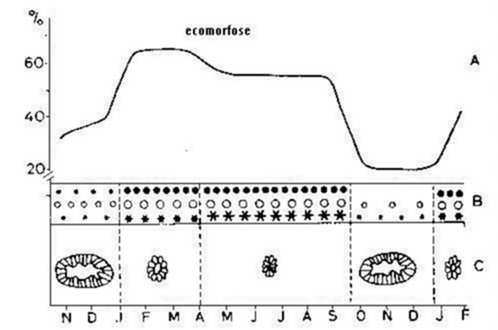
Fig. 3.4 Schema van de seizoengebonden cyclus van het verschijnsel ecomorfose bij Hypogastrura tullbergi (naar Lauga-Reyrel, 1979)
A: Volume vetlichaampjes (%) van totaal lichaamsvolume; B: Ontwikkeling van de vetcellen; C: Veranderingn in middendarmstructuur;
(○) Eiwitbestanddelen; (●) vetlichaampjes; (*) uraat (zout van urinezuur) cellen
De morfologische veranderingen doen zich vooral ter hoogte van de cuticula voor. Deze verdikt zich en gaat een aantal kenmerkende abdominale stekels ("abdominal spines") vertonen. Een regressie van de furca en de monddelen wordt meestal ook waargenomen (Casagnau, 1956; Najt, 1983). Gelijktijdig gaat het volume van het vetlichaam toenemen, tot het in uitzonderlijke gevallen tot 70% van het lichaamsvolume inneemt (Joosse, 1983)(fig. 3.4). De dieren migreren diep in de bodem. Ze nemen gedurende de ganse tijdsduur van de ecomorfose geen voedsel op en blijven inactief. Er is een scherpe daling van het gehalte aan vrije aminozuren in de haemolymfe (Dalens, 1982), terwijl de waterregulatie slechter wordt (Dalens & Vannier, 1979). Er vindt met name een omschakeling plaats van een mesofiel type evaporatieflux naar een hygrofiel type. Dit wordt echter gecompenseerd door een verbeterde cuticulaire weerstand (verdikken cuticula, verminderde lichaamsoppervlakte), die een betere barrière vormt tegen waterverlies door evaporatie (Dalens & Vannier, 1983). In tegestelling met soorten die in anhydrobiose gaan, is ecomorfose niet onmiddellijk omkeerbaar (> 15 dagen) (Cassagnau, 1956).
3.9.1.3 Adaptaties aan langdurige submersie
Kortstondige submersie van springstaarten heeft meestal geen enkele invloed op de dieren. De aanwezigheid van een luchtlaag rond het lichaam zorgt er immers voor dat er geen zuurstofgebrek optreedt. Bij Anurida maritima, een bewoner van slikken en schorren, is submersie zelf een onderdeel van de gewone dagcyclus (Joosse,1966). Hypogastrura viatica is zelf in staat om de ganse levenscyclus onder water door te maken (Mertens et al.,1983).
Bij langdurige submersie neemt de beschikbare hoeveelheid zuurstof in de capillairen meestal snel af (Zinkler,1983). Dit leidt bij de individuen, die niet opgespoeld worden en in de capillairen gevangen zitten, uiteindelijk tot de dood.
De eieren van springstaarten zijn daarentegen bijzonder resistent aan submersie (Marshall & Kevan, 1962). Tamm (1984) geeft als maximale submersieduur 309 dagen aan bij Isotoma viridis. Waarschijnlijk gaat het hier om een diapauze conditie (Berbiers, intern rapport). Deze bijzondere adaptatie laat springstaarten toe, om het gebied na droogvallen zeer vlug te herkoloniseren (Tamm, 1984).
3.9.1.4 Adaptaties aan negatieve temperaturen
Springstaarten zijn succesvolle kolonisatoren van antarctische, arctische en alpiene gebieden. In Antartica vertegenwoordigen ze 86% van alle gekende insektensoorten (Sømme, 1981). Het is dus evident dat de soorten die er voorkomen temperaturen onder het vriespunt kunnen verdragen. Ook springstaarten van gematigde klimaatzones moeten opgewassen zijn tegen zeer lage wintertemperaturen, te meer omdat in deze gebieden een isolerende sneeuwlaag vaak ontbreekt.
Bij invertebraten die aan temperaturen beneden het vriespunt van hun haemolymfe blootstaan, werden twee types van overlevingingsstrategie ontwikkeld: vorsttolerantie ("freezing tolerance") en vorstgevoeligheid ("freezing susceptibility")(Berbiers, intern rapport).
Vorsttolerante soorten vermijden vorstschade van de weefsels door een mechanisme te ontwikkelen van gecontroleerde extracellulaire ijsvorming bij lage temperaturen (-5 tot -12°C) (Block, 1981). Door de aanwezigheid van ijsvormende proteïnen gedurende de winter, wordt de mogelijkheid tot onderkoeling sterk gereduceerd. Al de tot nu toe onderzochte springstaarten zijn vorstgevoelig, maar Poinsot-Balaguer & Barra (1983) toonden aan dat Folsomides angularis in anhydrobiose in staat is om extreem lage temperaturen (-180°C) te doorstaan. Vorstgevoelige soorten kunnen de vorming van ijskristallen in de weefsels niet overleven (Zettel, 1984). Bevriezen wordt vermeden door in onderkoeling te gaan. Dit wordt bereikt door de concentratie van zogenaamde cryoprotectantia in de haemolymfe te verhogen (glycerol, ribitol, sorbitol, enz.). Hierdoor wordt het vriespunt van de haemolymfe sterk verlaagd (max. tot -38°C, SPC of "supercoolingpoint") (Sømme & Conradi-Larsen, 1977). De aanwezigheid van vrij water in de weefsels wordt hierbij sterk beperkt.
Het onderkoelingsbereik bij springstaarten wordt nog vergroot door het ledigen van de darm (fig.3.5). Algemeen wordt aangenomen dat in de darminhoud elementen voorkomen die als nucleus voor ijsvorming optreden (Salt, 1968). Block & Young (1978) demonstreerden dat een contaminantenvrij dieet een maximaal onderkoelingsbereik verzekert.
Ondanks het in onderkoeling gaan, blijft het gevaar voor contactbevriezing ("inoculative freezing") bij bepaalde springstaarten bijzonder groot (Berbiers, internrapport). Contactbevriezing treedt op wanneer door een dampdrukverschil tussen de onderkoelde haemolymfe en het omringende ijs, een uitwaartste stroom van waterdamp optreedt. Deze waterdamp bevriest op het lichaam en groeit door de transcuticulaire kanalen naar binnen aan, tot het ijsfront contact maakt met de haemolymfe. Op dit ogenblik lijkt spontane bevriezing onafwendbaar. Springstaarten actief bij temperaturen lager dan het vriespunt van hun haemolymfe, winteractieve springstaarten die vaak in contact komen met ijs of sneeuw en overwinterende, inactieve springstaarten die niet volledig beschermd zijn tegen het insijpelen van vocht lopen allen het risico van contactbevriezing.
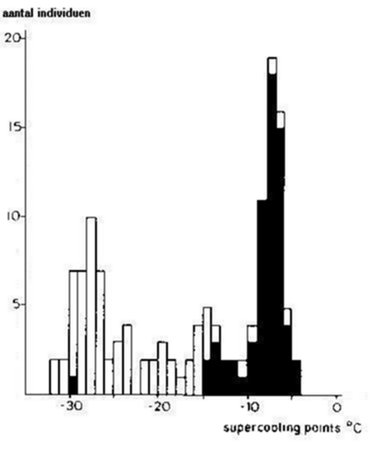
Fig. 3.5 "Supercoolingpoints" (°C) gerelateerd aan de darminhoud bij Tetracanthella wahlgreni.
■ = met darminhoud, □ = zonder darminhoud (naar Sømme & Conradi-Larsen, 1977).
< <
Zettel (1984) beschreef bij springstaarten proteïnen die een uitgesproken thermale hysteresis veroorzaken, m.a.w., het vriespunt van de haemolymfe wordt veel lager dan het smeltpunt ervan. IJskristallen die door contactbevriezing in het lichaam worden gevormd, worden door de thermale hysteresis proteïnen (THP) ingekapseld. Hierdoor wordt verder aangroeien van de ijsnucleus vermeden. Ook polyolen zorgen ervoor dat contactbevriezing niet kan optreden. In de Zwitserse vooralpen overwintert Entomobrya nivalis in een inactieve toestand, verscholen onder de bast van sparren. Bij deze soort is er dan ook een grote concentratie aan THP’s en polyoververzadigde alcoholen aanwezig. Zettel & Meier (1997) toonden aan dat de polyolen enkel aanwezig zijn tijdens de inactieve fase, terwijl THP’s bijkomende bescherming bieden gedurende de overgangsfases herfst-winter, winter-lente, wanneer de dieren wel nog actief zijn maar vorst reeds kan optreden. Experimenten betreffende het op gang brengen van deze anti-vries synthese gaven temperatuur en fotoperiode als aanwijzende factoren aan (Zettel & Meier, 1997).
Naast deze fysische aanpassingen om bevriezing te vermijden, vertonen springstaarten die in besneeuwde habitatten voorkomen ook veranderingen in hun gedragspatroon. Ze vluchten van het sneeuwoppervlak naar de warmere omgeving onder het sneeuwpakket (Sømme, 1976). Verscheidene soorten migreren op- en neerwaarts in de sneeuw waarbij deze locomotie gereguleerd wordt door de temperatuur of inkomende zonnestraling (Leinaas,1981; Fox & Stoud, 1986).
Vervolgens hebben zich ook reeds morfologische aanpassingen ontwikkeld, cyclomorfose genaamd (Leinaas, 1981; Fjellberg, 1976, 1978). Bij Hypogastrura lapponicus bijvoorbeeld verschijnen bij de aanzet van de winter opvallende tandjes op de dens van de furca (fig.3.6). Ze worden beschouwd als een aanpassing aan de springactiviteit op het sneeuwoppervlak.
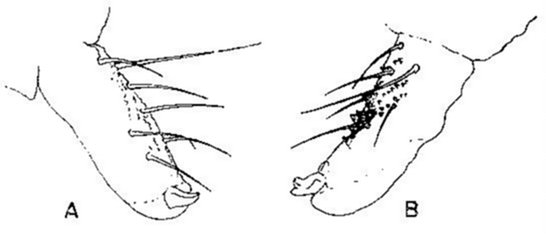
Fig. 3.6 Cyclomorfose in de furca van Hypogastrura lapponicus.
A: Zomervorm. B: Wintervorm met tandjes (naar Leinaas, 1981)
Ook springstaartsoorten van gematigde streken zijn metabolisch voorbereid op de koude winter. Individuen van verschillende soorten vertonen een veranderende tolerantie voor lage temperaturen (fig.3.7).
Een temperatuur waarbij nog leven mogelijk is tijdens de winter, is vaak lethaal tijdens de zomer. Testerinck (1982) beschreef een vorm van diapauze (oligopauze) om de koude periode te overbruggen: bij de aanzet van de winter kan een reductie van de opgestapelde energie opgemerkt worden.
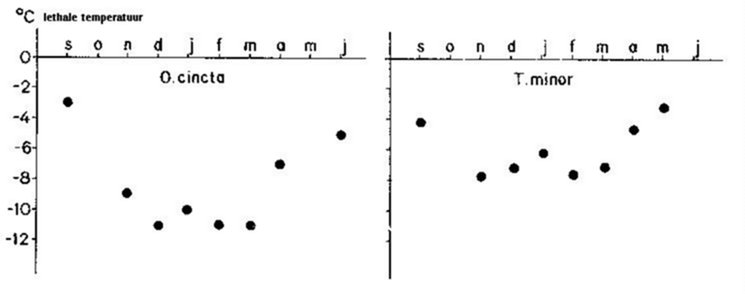
Fig. 3.7 Seizoengebonden variatie in lethale temperatuur bij Orchesella cincta en Tomocerus minor (naar Van Der Woude).
In figuur 3.8 is te zien dat de cirkel van de herfst smaller is. Verder is ook de energieverdeling verschillend in de lente en de herfst: naar de winter toe wordt meer energie geïnvesteerd in het behoud van het metabolisme en de reproduktie wordt volledig stopgezet. Deze strategie zou afwezig zijn in dieper levende soorten (Verhoef & Li, 1983).
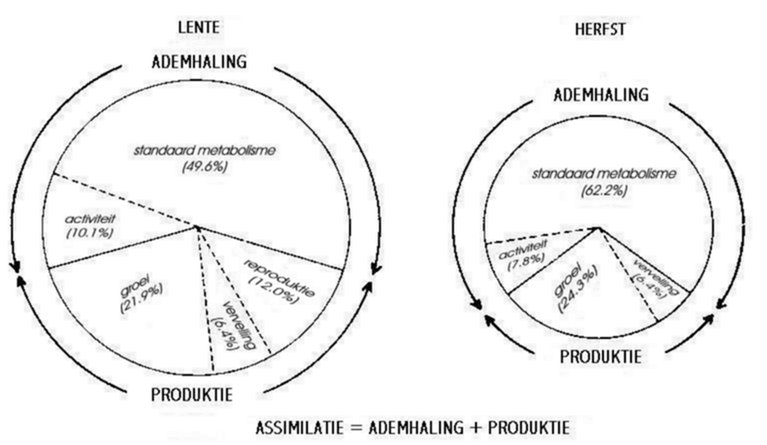
Fig. 3.8 Verdeling van de opgestapelde enrgie (%) over de verschillende activiteiten van Orchesella cincta in de lente en herfst.
Go to the
first,
previous,
next,
last
chapter,
overview.