
5. Materiaal en methode
5.1 Bemonsteringsmethodes
Afhankelijk van de verschillende doelstellingen van deze licentiaatsverhandeling, wordt de meest relevante bemonsteringsmethode gehanteerd.
5.1.1 Bepaling van de locomotorische activiteit
De locomotorische activiteit van de collembolen wordt bestudeerd door gebruik te maken van de pitfall-methode. Hemiëdafische, maar ook epigeïsche springstaarten kunnen gemakkelijk gevangen worden met behulp van pitfalls. De vangopbrengst van de springstaarten wordt niet alleen beïnvloed door hun locomotorisch activiteit en densiteit, maar ook door de efficiëntie van de val zelf (Adis, 1979). Zoals ook door Joosse & Kapteyn (1968) is weergegeven, zijn de vangopbrengsten niet noodzakelijk een correcte weergave van de normale locomotorische activiteit van de verzamelde soorten. Ze hebben aangetoond dat het ingraven van de pitfalls een locale stijging van het CO2-gehalte in de bodem veroorzaakt. Sommige collembolensoorten reageren hierop met een sterk verhoogde activiteit. Deze beïnvloeding van de locomotorische activiteit is niet voor alle soorten gelijk en niet constant in de tijd. Dit "digging-in effect" bemoeilijkt dus een vergelijking tussen verschillende staalreeksen (Berbiers et al., intern rapport). Naast dit "digging-in effect" kunnen ook verscheidene biotische en abiotische factoren de locomotorische activiteit beïnvloeden (Joosse & Groen,1970; Joosse & Testerink, 1977; Whitford et. al., 1981; Fox & Stoud, 1986).
In het huidige onderzoek gebruiken we pitfalls met een diameter van 14 mm. Uit een evaluatie van enkele bemonsteringsmethodes voor het bestuderen van collembolen blijkt dat een reductie van de diameter van de pitfalls de volgende voordelen biedt (Berbiers, intern rapport):
De drie habitatten (de duinroosruigte, de mosduinen in de voorduinen en de mosduinen langs de binnenduinrand) worden één of mogelijks twee keer per maand bemonsterd gedurende één jaar (1/5/99-18/4/00). De ledigingsdata zijn terug te vinden in tabel 5.1.
Tabel 5.1 Ledigingsdata van staalname
|
15/05/1999 |
26/10/1999 |
|
16/06/1999 |
14/11/1999 |
|
03/07/1999 |
01/12/1999 |
|
08/07/1999 |
06/01/2000 |
|
02/08/1999 |
05/02/2000 |
|
13/08/1999 |
29/03/2000 |
|
09/09/1999 |
18/04/2000 |
|
29/09/1999 |
|
Op deze data worden de buisjes verzameld en vervangen door een nieuwe reeks. Verscheidene stalen van het mosduinhabitat zijn onbruikbaar wegens overstuiving. In de binnenduinen en de duinroosruigte zijn verschillende bodemvallen weggenomen. Dit leidt tot een gevoelige tekortkoming van de data. Er zijn namelijk geen gegevens beschikbaar van de staalname in de mosduinen van 14/11/99 tot 1/12/99 en van de bemonstering in de binnenduinrand in de periode van 5/2/00 tot 9/3/00. De periode van 26/10/99 tot 14/11/99 levert geen enkel resultaat op daar alle buisjes verdwenen zijn. Daarnaast zijn in het laboratorium nog een aantal buisjes stuk gesprongen, met uitdroging van de organismen tot gevolg. Dit is hoogstwaarschijnlijk veroorzaakt door een te grote druk van het afsluitmechanisme in combinatie met een hogere temperatuur. Dit verlies aan vangstgegevens brengt wel enige consequenties met zich mee voor de verwerking van de resultaten.
Het principe van de pitfall-methode is eenvoudig: per habitat worden tien glazen proefbuisjes (Ø 14 mm, diepte 65 mm) lineair in de bodem geplaatst, enkele milimeters onder het bodemoppervlak, met een onderlinge afstand van ongeveer 30 cm. De buisjes zijn gevuld met een 4% formaldehyde-oplossing om de gevangen bodemaktieve organismen direct te fixeren. Om de oppervlaktespanning te verminderen wordt daarenboven een scheutje detergent toegevoegd . Dit voorkomt dat sommige specimens weer uit de pitfall kruipen. Er wordt een 4% formaldehyde-oplossing gebruikt daar dit niet zo snel verdampt als alcohol, maar het toxisch etiket is dan weer een negatief punt.
In het laboratorium wordt de formaldehyde-oplossing verwijderd door middel van een filter (200-300 µm) zodat de organismen deels in het buisje en deels in de filter achterblijven. Vervolgens worden beiden nagespoeld met alcohol en de hele fractie wordt in een petrischaal opgevangen.
De populatiestructuur van de meest abundante springstaartsoort zal bepaald worden aan de hand van individuen gevangen met deze methode.
5.1.2 Verticale verspreiding
Dit onderzoek wordt uitgevoerd in het mosduinhabitat van de voorduinen. De springstaarten die gerelateerd zijn aan het bodemoppervlak worden verzameld met behulp van een zuigtoestel of aspirator, terwijl de diepere bodemlagen bemonsterd worden aan de hand van bodemstalen.
Met de aspirator wordt een vooropgestelde oppervlakte van (40 × 40) cm2 bemonsterd. Het opgezogen materiaal (zand, plantenmateriaal, dierlijke organismen, etc.) wordt overgebracht in een pot die een 4% formaldehyde-oplossing bevat. De bovenste bodemlagen kunnen bemonsterd worden door gebruik te maken van een stalen cilinder met een diameter van 8 cm en een lengte van ± 20 cm. Een staalname van de diepere bodemlagen is mogelijk met behulp van een gutsboor. Dit is een 50 cm lange, halfopen smalle (Ø 3 cm) cilinder (fig.5.1).
Fig. 5.1 A) stalen cilinder. B) gutsboor.
Het nemen van een bodemstaal bestaat erin, de stalen cilinder met de scherpe rand in de bodem te duwen door middel van een draaiende beweging. Na het optillen van de gevulde cilinder kan het staal verdeeld worden in substalen. Bij het gebruik van een gutsboor kan een directe onderverdeling gemaakt worden. Wanneer de gesloten cilinder gebruikt wordt, moet de inhoud eerst zo volledig mogelijk verwijderd worden. Pas daarna kan de bovenste bodemlaag (vegetatie- en fragmentatielaag) gescheiden worden van de daaronderliggende zandlaag. Het verzamelde materiaal wordt gefixeerd en in het labo wordt de aanwezige pedofauna geëxtraheerd.
Het bodemmateriaal wordt in een spaghettizeef gebracht waaronder zich een maatkolf bevindt. Zo worden grovere deeltjes, zoals takjes en mosfragmenten gescheiden van de restfractie. Deze restfractie bevat nog aanzienlijke hoeveelheden zand die door herhaalde draaibewegingen, gevolgd door decantatie verwijderd worden. Als gevolg van de rotatiebeweging komt het organisch materiaal aan de oppervlakte drijven en kan het overgegoten worden in twee vertikaal opgestelde zeven met een maaswijdte van respectievelijk 2 mm en 200 µm. De onderste zeef, die de restfractie met de gewenste fauna bevat, wordt nagespoeld met alcolhol zodat alle aanwezige organismen verzameld worden.
De staalname vond plaats op het einde van de zomer (9/9/99) en in de lente van het daaropvolgend jaar (15/5/00). Van de tien bodemstalen, verzameld op 9/9/99 werden enkele substalen genomen en gecontroleerd op de aanwezigheid van springstaarten, met een negatief resultaat als gevolg. De helft van deze stalen was genomen met de gutsboor en ook hier konden geen collembolen aangetroffen worden. De procedure voor de bemonstering in de lente werd dan enigszins veranderd. De staalname diende zowel ‘s namiddag als in de late avond uitgevoerd te worden zodat een vergelijking kon gemaakt worden tussen de resultaten. Aanvankelijk was de datum van de bemonstering tijdens de lente 18/4/00, maar na een fixatiefout, met schimmelvorming tot gevolg, moest de staalname opnieuw uitgevoerd worden. Ook deze keer (8/5/00) werden geen resultaten binnengehaald; en dit dankzij het noodlottige regenweer in België. Door de opgeloopte vertraging en het daaruit voortvloeiende tijdsgebrek werd bij een laatste en succesvolle poging op 15/5/00 het aantal stalen sterk gereduceerd. Deze reductie bestond erin dat in plaats van tien, slechts vijf kwadranten (40 × 40) cm2 bemonsterd werden en dat het aantal bodemstalen herleid werd van tien naar drie. De staalname tijdens de namiddag verliep van 16u.50 tot 18u.40. ‘s Avonds om 22u.15 werd het hele protocol herhaald.
5.1.3 Horizontale verspreiding
Zoals de locomotorische activiteit wordt ook het voorkomen van aggregaten aan de hand van pitfalls bestudeerd. Ze hebben een diameter van 34 mm en een lengte van 68 mm. De bemonstering is éénmalig (9/3/99-29/3/99) en wordt uitgevoerd op de overgang van de mosduinvlakte naar de duinroosruigte. In totaal worden 40 bodemvallen kruisgewijs opgesteld in vijf verschillende microhabitatten (A - E). Habitatten A tot en met C bevatten elk tien pitfalls, terwijl D en E er slechts vijf bezitten. Uit de verzamelde gegevens zal ook de predator-prooi relatie nagegaan worden. Voor elk microhabitat worden metingen uitgevoerd omtrent de strooisellaag, de fragmentatielaag (F-laag) en de bodemontwikkeling (B-O). Daarnaast wordt een korte typering gemaakt van het vegetatiepatroon (tabel 5.2).
Tabel 5.2 Overzichtstabel van de bodemkenmerken en vegetatietypes van de microhabitatten (A-E).
|
|
A |
B |
C |
D |
E |
|
vegetatietype |
duingrasland |
kruidenvegetatie |
open mosduin |
open mosduin |
gras/mosduin |
|
% mosbedekking |
50 |
5 |
60 - 70 |
60 - 70 |
50 |
|
strooisellaag (+/-) |
- |
+ |
+ |
- |
+ |
|
fragmentatielaag (cm) |
1 |
1.5 - 2 |
0,3 |
0,3 |
1 |
|
bodemontwikkeling (cm) |
20 - 25 |
20 - 25 |
2 - 3 |
2 - 3 |
3 - 3.5 |
Habitat A:
Dit is een duingrasland dat voor de helft bestaat uit een moslaag met o.a. groot en klein duinsterretje (Tortula ruraliformis & T. calcicolens) en dikkopmos (Brachythecium rutabulum). Verder is ook veldbies (Luzula sp.) en fijn schapengras (Festuca filiformis) opvallend. Er is geen strooisellaag aanwezig en de fragmentatielaag beslaat slechts 1 cm van de 20 à 25 cm bodemontwikkeling.
Habitat B:
Dit habitat bestaat voornamelijk uit een kruidenvegetatie (3 cm) die het mos (5%) niet veel kansen biedt en de bodem bedekt met een stooisellaag. In het oog springend zijn o.a. grote tijm (Thymus pulegioides) en zonneroosje (Helianthenum nummelarium). De fragmentatielaag is 1,5 à 2 cm dik en de mate van bodemontwikkeling komt overeen met die van habitat A.
Habitat C:
Dit bevindt zich in een open mosduinvlakte waar 60 tot 70% van het oppervlak bedekt is met mos. De aanwezigheid van een kruipwilg (Salix repens) is verantwoordelijk voor het voorkomen van een strooisellaag. De fragmentatielaag is uiterst dun (3 mm) en ook de bodemontwikkeling is minimaal (2-3 cm).
Habitat D:
Dit habitat is identiek met het vorige, met als enige verschil dat er geen kruipwilg (Salix repens) en dus geen strooisellaag voorkomt.
Habitat E:
Hier bestaat de vegetatie zowel uit mos (50%), grassen (40%) en kruiden (10%). Naast rood zwenkgras (Festuca rubra s.l.), duinreigersbek (Erodium cicutarium) en sint-janskruid (Hyperium perforatum L.), is ook kruipend stalkruid (Ononis repens) goed vertegenwoordigd. Het afsterven van de kruiden zorgt ervoor dat de mate van bodemontwikkeling iets groter is dan in habitat C en D (3-3,5 cm). De fragmentatielaag heeft een dikte van 1 cm en er is een strooisellaag aanwezig.
5.2 Determinatie en metingen
5.2.1 Determinatie
De determinatie werd uitgevoerd met behulp van een Wild M3 en een Wild M5 stereomicroscoop en een Kyowa medilux microscoop. De identificatie tot op soortniveau (waar mogelijk) gebeurde volgens Gisin (1960). Het determineren werd enigszins vergemakkelijkt omdat we reeds een idee hadden welke soorten er in een duinecosysteem kunnen voorkomen. Een samenvattende genustabel met de mogelijke voorkomende soorten, opgesteld door Dries Bonte (intern rapport), was bijzonder handig. Moeilijk te determineren soorten werden door Frans Janssens (collembologist) geïdentificeerd (persoonlijke communicatie). Toch verliep de determinatie niet zonder problemen. Ten eerste bleven beschadigde soorten en zeer kleine juvenielen vaak onbenoemd. Vervolgens heeft het grote verspreidingsgebied van de meeste springstaarten tot gevolg dat er binnen één soort heel wat variatie kan optreden. Wallace (1973) bijvoorbeeld splitse een grote verzameling van Sminthurus viridis op in drie soorten, gebaseerd op hun kleurenpatroon. Het gaat om Sminthurus viridis, sminthurus marmoratus en Sminthurus nigromaculatus. Niettegenstaande dat de drie soorten morfologisch gelijkaardig zijn (en moeilijk te scheiden na het vervagen van de kleuren onder invloed van het fixatief), vertonen ze duidelijke verschillen in de geografische gebieden die ze bezetten. Deze specimens worden beschouwd als aparte soorten, behorende tot dezelfde morfologisch homogene "species-groep" (Hopkin, 1997). Tenslotte leidt het gebrek aan type-specimens en de vaak dubbelzinnige beschrijvingen van vroeger tot opmerkelijke taxonomische verwarring. In de periode voordat de telefoon of een betrouwbare postdienst bestond, waren de meesten er zich niet van bewust dat de soort die ze zopas benoemd hadden, reeds elders beschreven was. Lubbock (1873) besteedde in het bijzonder aandacht aan de synoniemen die oprezen, als gevolg van een gelijktijdige publicatie van Bourlet (1839, 1843) en Nicolet (1847). Het grootste gedeelte van de synoniemen voor de genera zijn ondertussen opgehelderd (Ellis & Bellinger, 1973, 1984). Voor de soorten blijft het nog steeds iets moeilijker. Nog steeds vormt het voorhanden zijn van talrijke synoniemen en struikelblok voor vele systematici en taxonomen. Fjellberg (2000, email correspondentie) is bezig met het opstellen van een werk dat ons een beter zicht zou moeten geven op het zogenaamde Isotoma viridis complex. Hij ontdekte het bestaan van twee wijdverspreide soorten, verscholen onder dezelfde naam Isotoma anglicana. Welk specimen het originele is dat Lubbock in 1862 beschreef, weet niemand. Waarschijnlijk zullen in de toekomst, met de vooruitgang van moleculaire methodes voor determinatie, nog meer gevallen aan het licht komen (Fjellberg, 2000, email correspondentie). Een redenering volgens het principe van Gause2, namelijk, dat bij eenzelfde nichegebruik het waarschijnlijk om één soort gaat, daar er anders competitie zou optreden, kan ons soms heel wat tijd besparen bij de determinaties.
In totaal werden 18 soorten geïdentificeerd en geteld. Om een tijdrovende determinatie te vermijden, werden bij het tellen alle Sminthuridae samen genomen. Het was echter niet de bedoeling een intensieve inventarisatie op te stellen maar een fenologische studie uit te voeren.
5.2.2 Lengtemetingen van Isotoma viridis
Om inzicht te verwerven in de populatiestructuur van de meest abundante springstaartsoort, namelijk Isotoma viridis, zijn metingen van de lichaamslengte noodzakelijk. De populatie, verzameld met de kleine pitfalls in de duinroosruigte leek het meest geschikt voor deze studie. Van elke periode werden willekeurig 30 individuen (indien mogelijk) gemeten volgens de methode van Tamura (1974).
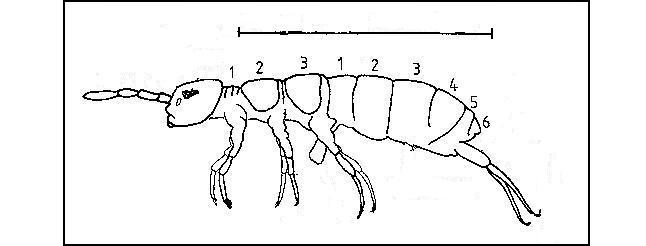
Fig. 5.2 Aanduiding van de gemeten lichaamslengte.
5.2.2.1 Beschrijving van de methode
Specimens van Isotoma viridis, gefixeerd in een 4% formaldehyde-oplossing en bewaard in 70% ethanol, worden in een petrischaal op de voorwerptafel van een stereomicroscoop (Wild M5) gebracht, voorzien van een tekenspiegel. Op een tekenblad wordt met behulp van deze tekenspiegel een scherp omlijnde afbeelding van het specimen verkregen. De standaardlichaamslengte van de collembolen kan zo nauwkeurig doorgetekend worden als een gebogen lijn vanaf het tweede thoracale tot het laatste abdominale segment in lateraal zicht (fig.5.2). De lengte van de getekende krommen wordt met behulp van een "Summa Sketch III Professional" digitaliseertablet berekend, met een nauwkeurigheid van 0,01 mm.
5.2.2.2 Effect van het fixatief
Uit een intern rapport blijkt dat een fixatie in een 4% formaldehyde-oplossing bij Isotoma viridis een extensie van ongeveer 13% veroorzaakt. Het effect van het fixatief is constant bij individuen van verschillende lengte. Mertens et al. (1982) toonde aan dat fixatie in picrinezuur en een bewaring in 70% ethanol een verlenging van 10% veroorzaakt bij Orchesella cincta.
5.2.2.3 Relatie tussen standaardlengte en totale lengte
De resultaten van Berbiers (intern rapport) geven weer dat de standaardlichaamslengte bij Isotoma viridis ongeveer 80% van de totale lichaamslengte bedraagt bij kleine individuen. Deze ratio blijft echter niet constant zodat grotere springstaarten een relatief langer lichaam (standaardlengte) hebben.
5.3 Statistische verwerking
De statistische verwerkingen zijn uitgevoerd met behulp van het statistisch software pakket (Statfort, 1998). Voor de grafische voorstelling van de populatiestructuur van Isotoma viridis is het programma Sigmaplot (SPSS, 1997) gebruikt.
5.3.1 Parametrische variantie-analyse: ANOVA = ANalysis Of VAriance
De doelstelling van een variantie-analyse is om de homogeniteit of heterogeniteit van groepsgemiddelden met elkaar te vergelijken. De rekenkundige methode van de analyse bestaat erin om de som der kwadraten (s.s.:sum of squares) en de vrijheidsgraden (d.f.: degrees of freedom) op te splitsen, eerder dan de varianties direct te analyseren. De varianties worden vergeleken aan de hand van de F-test en er wordt een significantieniveau van 5% (p=0.05) gebruikt.
De variantie-analyse wordt voornamelijk gebruikt bij de beschrijving van het locomotorisch activiteitspatroon van de springstaarten. De analyse wordt toegepast op de volgende variabele selectiecriteria: de periode en het habitat. Er wordt ook bepaald of een combinatie van beide variabelen (interactie) een significant verschil oplevert. Omdat de verschillende periodes niet altijd evenveel dagen bevatten, wordt het aantal gevangen specimens per staal gedeeld door het totaal aantal dagen van de bemonsteringsperiode. De variantie-analyse wordt uitgevoerd op deze gestandariseerde aantallen, die het gemiddeld aantal springstaarten per dag weergeven. Vervolgens wordt ook in het onderzoek naar de horizontale verspreidng een variantie-analyse uitgevoerd. Dit om na te gaan of er significante vershillen optreden in het relatief voorkomen van de collembolen in de verschillende microhabitatten.
5.3.2 Niet-geplande vergelijkingen of a posteriori testen (post-hoc test)
Wanneer de F-test bij de variantie-analyse wijst op een significant verschil tussen de gemiddelden kunnen daarenboven niet-geplande vergelijkingen uitgevoerd worden tussen de gemiddelden. Opvallende verschillen tussen twee gemiddelden kunnen getest worden op hun significantie. Deze testen worden uitgevoerd met de methode van Scheffé, die gebaseerd is op de F-verdeling. De resultaten kunnen aan de hand van een boxplot grafisch voorgesteld worden.
5.3.3 Niet-parametrische variantie-analyse: Mann Whitney U test
Hier wordt de significantie van het verschil tussen twee onafhankelijke groepen bekeken. Deze test is een test van rangordes, dit wil zeggen dat de tellingen vervangen worden door hun rangorde. De Mann Whitney U test wordt toegepast in het onderzoek naar de verticale verspreiding van de collembolen, met de periode (dag-nacht) als onafhankelijke variabele. De significantie van de temperatuur- en luchtvochtigheidsverschillen wordt eveneens met deze test onderzocht. Opnieuw kan een grafische voorstelling verkregen worden aan de hand van een box-whisker plot.
5.3.4 ‘Spearman-rank’ correlatiecoëfficiënt
Deze niet-parametrische test wordt gebruikt om de mate van samenhang te bepalen tussen twee variabelen. De methode is gebaseerd op het toekennen van rangordes aan de verschillende data. De correlatiecoëfficiënt wordt verwacht gelijk te zijn aan –1 of +1 bij perfecte negatieve of positieve samenhang en gelijk aan 0 bij volledige afwezigheid van samenhang. De correlatie tussen het voorkomen van de springstaarten en de verschillende bodemkenmerken van de microhabitatten en de associatie tussen de activiteit van spinnen en collembolen wordt met deze test onderzocht. De resultaten worden voorgesteld in een scatterdiagram.
5.4 Karakterisatie van de stations (habitatten)
De situering van de drie stations voor de studie naar de locomotorische activiteit werd zo gekozen dat een vergelijking kan gemaakt worden tussen enerzijds verschillende vegetatietypes en anderzijds klimatologische factoren zoals een verschillende luchtvochtigheidsgraad (afhankelijk van de afstand tot de zee). De drie stations werden bemonsterd gedurende een periode van 1/5/99 tot 18/4/2000.
Station MD: droog Tortula ruralis ssp. ruraliformis mosduin
Dit station is gelegen in de chaotische voorduinen. Hier is geen hoge kruid- of struweellaag aanwezig. De vegetatie wordt volledig gedomineerd door een dichte, lage moslaag voornamelijk bestaande uit groot en klein duinsterretje (Tortula ruraliformis en T. calcicolens). Begeleidende therofyten zijn o.a. vroegeling (Erophila verna), vroege haver (Aira praecox), duinreigersbek (Erodium cicutarium ssp. dunense), zachte ooievaarsbek (Geranium molle), zandhoornbloem (Cerastium semidecandrum), rood zwenkgras (Festuca rubra s.l.), zanddoddegras (Phleum arenarium) en muurpeper (Sedum acre). Aan de zeezijde vormt kruipend stalkruid (Ononis repens) de overgang met de helmduinen van de zeereep, terwijl in het zuiden duinroos (Rosa pimpinellifolia) de duinmosvlakte begint binnen te dringen.
Station DR: noordelijke helling van paraboolarm met duinroos (Rosa pimpinellifolia)
Hier is duinroos (Rosa pimpinellifolia) zeer dominant en vormt een dichte vegetatie met weinig ondergroei. Van de weinig aanwezige andere hogere planten zijn zachte dravik (Bromus hordeaceus), zonneroosje (Helianthemum nummelarium), fijn schapengras (Festuca filiformis) en grote tijm (Thymus pulegioides) opvallend aanwezig. Dit vegetatietype is maximum 40 à 50 cm hoog en vertoont praktisch geen strooisel.
Station BD: matig droog Tortula ruralis ssp.ruraliformis mosduin
Dit station situeert zich in de relatief stabiele zuidelijke duinenrij ten noorden van de aangeplante bomen langs de binnenduinrand. Het vegetatiepatroon is gelijkaardig aan dat van station MD.
Go to the
first,
previous,
next,
last
chapter,
overview.