
6. Resultaten
6.1 Locomotorische activiteit
6.1.1 Globale resultaten
In totaal werden meer dan 11600 springstaarten gevangen, behorende tot 18 verschillende soorten. Tabel B.1 (B=bijlage) geeft een overzicht van de ruwe data. De zes meest abundante soorten worden weergegeven in tabel 6.1. Het betreft de volgende soorten: Isotoma viridis, Xenylla maritima, Entomobrya nivalis, Folsomia cf. quadrioculata, Crytopygus thermophilus en Orchesella cincta. De Sminthuridae worden uiteraard buiten beschouwing gelaten, daar het hier niet om een soort gaat, maar om een familie. De soorten behorende tot deze familie zijn vaak zeer klein en daarom moeilijk te determineren. De verdere verwerking zal hoofdzakelijk betrekking hebben op Isotoma viridis, Xenylla maritima en Entomobrya nivalis.
Tabel 6.1 De zes meest abundante soorten, met het
gemiddeld aantal specimens per staal.
6.1.2.1 Isotoma viridis (Bourlet, 1839)

Fig.6.1 Isotoma viridis (x 6).
Isotoma viridis is een middelgrote (maximale lengte 4,3 mm) hemiëdafische (soms epigeïsche) springstaart, behorende tot de familie Isotomidae. Deze soort is bijzonder variabel in kleur, heeft een goed ontwikkelde furca en antennes met donkere uiteinden. De lengte van het derde en vierde abdominaal segment vertoont geen noemenswaardig verschil. Het is een hygrofiele soort die in Europa voorkomt vanaf de Arktische eilanden tot Spanje (Gisin,1960) en ook in Canada te vinden is (Aitchison, 1984). Van deze soort is bekend dat ze winteractief is en zelfs op de sneeuwlaag kan voorkomen (Aitchison, 1983).
6.1.2.2 Entomobrya nivalis (Linné,1758)
Fig 6.2 Entomobrya nivalis (x 6)
Entomobrya nivalis, behorende tot de familie Entomobryidae, is gekenmerkt door een lichaam zonder schubben, met borstels en een typische pigmentering. De lichaamslengte varieert van 0,5 tot 2,18 mm. Het vierde abdominaal segment is meestal veel langer dan het derde. Het is een hemiëdafische soort die men ook kan aantreffen onder de bast van naaldbomen. Deze soort is verspreid over gans Europa en Noord-Amerika. Van Entomobrya nivalis is geweten dat ze in een inactieve toestand de koude winter kan doorbrengen (Zettel & Meier, 1995).
6.1.2.3 Xenylla maritima (Tullberg, 1869)
Xenylla maritima is een typische xerofiele soort die ook op boomstammen en in naaldstrooisel te vinden is (Gisin,1960). Ze heeft een maximale lengte van 1,4 mm. Zoals de twee vorige soorten, komt ook deze voor over geheel Europa. Ze heeft een paars-donkerblauwe pigmentering, wederzijds vijf ogen, een gereduceerde furca en korte antennes. Postantennaal orgaan en empodium ontbreken. Deze soort behoort tot de familie Poduridae en vertoont geen winteractiviteit (Aitchison, 1984).
6.1.3 Bespreking locomotorische activiteit
Van periode 10 (14/11/99-1/12/99) en 13 (5/2/00-9/3/00) zijn respectievelijk geen gegevens beschikbaar van de staalname in de mosduinen en de binnenduinrand wehens overstuiving en wegname, zodat de data van deze periodes helaas niet gebruikt kunnen worden.
De vangstaantallen in tabel B.1 geven aan dat Cryptopygus thermophilus bijna uitsluitend voorkomt in de mosduinen en dan voornamelijk in de voorduinen vanaf maart tot eind juni. De sminthuridae blijken de hoogste activiteit te vertonen in september en oktober en dit uitsluitend in de duinroosruigte. Zeer opmerkelijk is de winteractiviteit van Folsomia cf. quadrioculata. De hoogste vangstaantallen worden waargenomen tijdens de maanden januari en februari, terwijl alle andere gevangen soorten dan juist weinig of geen activiteit vertonen. Deze soort is afwezig tijdens augustus en september en komt enkel voor in de duiroosvegetatie.
Voor de drie besproken soorten worden de volgende vraagstellingen getoetst:
6.1.3.1 Isotoma viridis
Tabel B.3, geeft aan dat er een significant verschil (p< 0.05) is tussen zowel de periodes (1), de habitatten (2), als de interactie tussen beiden (12).
Er wordt verwacht dat de soort gedurende bepaalde periodes van het jaar meer of minder actief zal zijn. Dit is precies wat we terugvinden in figuur 6.3. Isotoma viridis is vooral actief tijdens de lentemaanden maart, april, mei en juni, met een piek in de maand mei. Tabel B.4 geeft aan dat deze hogere aantallen significant zijn en de overeenkomstige box-whiskler plot is afgebeeld in figuur B.1. De rest van het jaar is de soort beduidend minder actief of minder aanwezig. Een dieptepunt wordt waargenomen in de maanden augustus en december. In de duinroosruigte is een duidelijk activiteitspatroon merkbaar: twee periodes van hogere locomotorische activiteit, van maart tot juli en in oktober en november, worden voorafgegaan door periodes van lage activiteit, van december tot februari en in augustus en september. Een overeenkomstig patroon werd vastgesteld door Berbiers et al.(intern rapport), tijdens een studie over de locomotorische activiteit van Isotoma viridis in een sterk begraasde weide in Melle. In de twee andere habitatten is de tweede periode van hogere locomotorische activiteit (oktober, november) helemaal niet zo uitgesproken.
Zoals tabel B.5 en figuur B.2 tonen, vertoont Isotoma viridis een gelijkaardig voorkomen in de mosduinen en de duinroos, maar is beduidend minder aanwezig in de binnenduinrand (p= 0.022; p= 0.002).
Uit tabel B.6 en figuur B.3 blijkt dat Isotoma viridis gedurende bijna het hele jaar homogeen voorkomt in de drie gebieden. Enkel in de eerste helft van de maand mei (1/5-15/5) is de soort opvallend meer vertegenwoordigd in de mosduinen.
6.1.3.2 Entomobrya nivalis
Ook hier vinden we een significant verschil (p=0.00) tussen de locomotorische activiteit in de verschillende periodes (1) en in de drie habitatten (2). Tevens is de activiteitsverdeling over de drie gebieden niet voor alle periodes gelijk (12)(tabel B.7).
Wanneer de verschillende habitatten buiten beschouwing worden gelaten, zien we op figuur B.4 dat enkel in het begin van de maand juli (periode 4) het aantal gevangen specimens beduidend hoger ligt (tabel B.8). Omdat Entomobrya nivalis uitsluitend in de duinroosruigte een representatieve locomotorische activiteit vertoont, zal enkel figuur 6.4A nader besproken worden. Deze soort vertoont duidelijk een verschillende activiteit in vergelijking met Isotoma viridis. We kunnen een lage activiteit vaststellen tijdens de wintermaanden december, januari en februari. Vanaf maart stijgt de activiteit, tot een piek bereikt wordt in juli, waarna ze geleidelijk aan opnieuw zal dalen. De piekwaarde is niet zo uitgesproken als bij Isotoma viridis. Wat nog opvallend is, is dat het activiteitsverschil tegengesteld is met dat van Isotoma viridis: wanneer Entomobrya nivalis een hoogtepunt bereikt in haar activiteit (in juli), is Isotoma viridis al bijna opnieuw aan een dieptepunt toe (in augustus). Tijdens oktober en november is een gelijkaardige activiteit waar te nemen.
Zoals reeds in figuur 6.4 te zien is, blijkt Entomobrya nivalis het duinrooshabitat te prefereren. Zowel tabel B.9 als figuur B.5 bevestigen dit.
Op figuur B.6 is te zien dat in een aantal periodes de locomotorische activiteit iets hoger ligt in de duinroos, maar enkel in de periode van 3/7 tot 8/7 is dit verschil significant (tabel B.10).
6.1.3.3 Xenylla maritima
Ook deze soort vertoont significante verschillen voor de drie onderzochte effecten (tabel B.11).
Terwijl de twee vorige soorten hun hoogste locomotorische activiteit vertonen tijdens de warmere dagen van de lente en het begin van de zomer, is deze soort het meest actief in de maanden september en oktober, wanneer het opnieuw al wat koeler wordt. Dit is duidelijk waar te nemen op figuur 6.5. Het hoge aantal individuen gevangen in het habitat langs de binnenduinrand, zorgt ervoor dat figuur B7 ook het begin van juli aangeeft als een periode met opvallend hogere activiteit. De significanties zijn weergegeven in tabel B12. Waarschijnlijk zijn hier, net zoals bij Isotoma viridis, twee periodes van hogere activiteit aanwezig. De meest uitgesproken periode is deze vanaf juni-juli tot het einde van november. De locomotorische activiteit gaat de hoogte in tot een piekwaarde bereikt wordt in september, waarna de activiteit opnieuw zal dalen en aanleiding geven tot een periode gekenmerkt door weinig of geen activiteit gedurende de maanden december, januari en februari. Daarna zal de locomotorische activiteit opnieuw een stijgende trend aannemen tot in april, terwijl deze in mei al opnieuw gedaald is. De waarden in deze laatste activiteitsperiode liggen opmerkelijk lager dan in de andere periode. In tabel B.12 en op figuur B.7 kunnen we zien dat in deze periode de activiteiten niet significant verschillen. In de mosduinen is een kleine verschuiving opgetreden; de hoogste activiteitswaarden worden hier niet verkregen in de maanden april en september, maar in maart en oktober. Xenylla maritima is afwezig in de mosduinen gedurende de maand augustus. De afwezigheid in november is een verkeerdelijke indruk die ontstaat omdat er geen data beschikbaar zijn van deze maand.
In tegenstelling tot wat de soortnaam laat vermoeden, vertoont Xenylla maritima een uitgesproken voorkeur voor het habitat in de binnenduinrand (tabel B.13, fig.B.8); maar als je weet dat het hier om een xerofiele soort gaat, is deze preferentie begrijpelijk.
Op figuur B.9 kunnen we zien dat de periodes met een significant hogere activiteit in de binnenduinrand, namelijk van 3/7 tot 8/7 en van 9/9 tot 26/10, voorafgegaan worden door periodes met een lichte, niet-significante activiteitsstijging (tabel B.14). Deze periode komt overeen met de eerder besproken activiteitsperiode van figuur 6.5.
6.1.4 Samenvatting
De drie soorten vertonen allen weinig of geen locomotorische activiteit tijdens de koude wintermaanden december, januari en februari. Xenylla maritima heeft een uitgesproken voorkeur voor de binnenduinrand, terwijl Isotoma viridis in dit habitat enkel tijdens de lente een waar te nemen activiteit vertoont. Entomobrya nivalis heeft haar thuishaven gevonden in de duinroosruigte. In tegenstelling tot de twee andere soorten, is deze gedurende één lange periode actief, met een hoogtepunt in de zomer. Bij Isotoma viridis is de hoogste activiteit waar te nemen in de lente en Xenylla maritima is het meest actief in de herfst. Deze habitatspreferentie is in bepaalde periodes wel meer uitgesproken, maar zal niet veranderen.
6.2 Populatiestructuur van Isotoma viridis
6.2.1 Bespreking
De populatie, verzameld met de kleine pitfalls in het duinrooshabitat leende zich het best voor dit onderzoek. Een studie van een natuurlijke populatie is hoofdzakelijk gebaseerd op metingen van de lichaamslengte. Na een indeling in lengteklassen, is het mogelijk een schatting te maken van de leeftijdssamenstelling van de populatie, verzameld gedurende één jaar (12 periodes), en inzicht te verwerven in de populatiestructuur van Isotoma viridis. Fenologische studies indiceren dat er één, twee of drie generaties bestaan gedurende een jaar, afhankelijk van de breedteligging van de bestudeerde populatie (Agrell, 1941; Milne, 1962; Joosse,1969; Weigmann, 1973; Fjellberg,1975; Aitchison, 1984).
Tabel 6.2 De periodes waarvan 30 individuen van Isotoma viridis gemeten zijn.
|
periode |
|
|
1 |
6/1/00-5/2/00 |
|
2 |
5/2/00-9/3/00 |
|
3 |
9/03/00-29/3/00 |
|
4 |
29/3/00-18/4/00 |
|
5 |
1/5/99-15/5/99 |
|
6 |
15/5/99-16/6/99 |
|
7 |
16/6/99-8/7/99 |
|
8 |
8/7/99-13/8/99 |
|
9 |
13/8/99-29/9/99 |
|
10 |
29/9/99-26/10/99 |
|
11 |
14/11/99-1/12/99 |
|
12 |
1/12/99-6/1/00 |
In totaal zijn 302 specimens gemeten met een lengte variërend van 0.338 mm tot 2.547 mm. Wanneer we de extensie van 13% ten gevolge van de fixatie in rekenschap brengen, bekomen we een lengte tussen 0.382 mm en 2.878 mm. De resultaten zijn terug te vinden in tabel B.15. De bekomen lengtes zijn verdeeld over 56 klassen met een klassebreedte van 0.05 mm, beginnende met de waarde 0.3 mm en eindigend met de waarde 3.0 mm. Tabel B.16 geeft het aantal collembolen per grootteklasse weer voor elke periode. In functie van de lichaamslengte is een frequentieverdeling opgesteld, welke te zien is in figuur 6.6. Het is niet uitgesloten dat de opeenvolgende pieken van de grafiek corresponderen met de instars. De eerste stijging van deze bimodale verdeling kan verklaard worden door het feit dat het hier gaat om pas uitgeslopen juvenielen, die nog niet veel aan levensbedreigende omstandigheden zijn blootgesteld. In functie van de toenemende ouderdom zal een daling van het aantal individuen optreden. Dit verklaart tevens de lage frequenties van de individuen groter dan 2.2 mm. De hoge reproductieve activiteit van de adulten is verantwoordelijk voor de frequentiestijging vanaf een lengte van 1.35 mm, daar de vangstopbrengst een functie is van de activiteit. Berbiers (intern rapport) toonde aan dat Isotoma viridis reproductief wordt bij het 10de instar. Bij het bereiken van dit instar, bedraagt de standaardlengte 1.30 (�0.17) mm. Globaal vertoont de groei van springstaarten een sigmoidaal verband. Dit houdt in dat er een geringere groeisnelheid is bij groter wordende individuen. De lage frequentie van de grootteklassen tussen 1.15 mm en 1.35 mm kan toegeschreven worden aan een aantal factoren. Allereerst komt dit lengte-interval overeen met de periode net voor het bereiken van de sexuele rijpheid. De individuen zijn wel actief, maar niet zo uitgesproken als in de reproductieve fase. Ten tweede wordt die periode gekenmerkt door een hoge groeisnelheid. Een ongeveer maandelijkse bemonstering kan dan al vlug enkele grootteklassen over het hoofd zien. Een derde factor is de reeds vermelde afname van het aantal individuen met toenemende leeftijd. De combinatie van een lage activiteit, een hoge groeisnelheid en een relatief lagere densiteit is verantwoordelijk voor het dieptepunt van de verdeling.
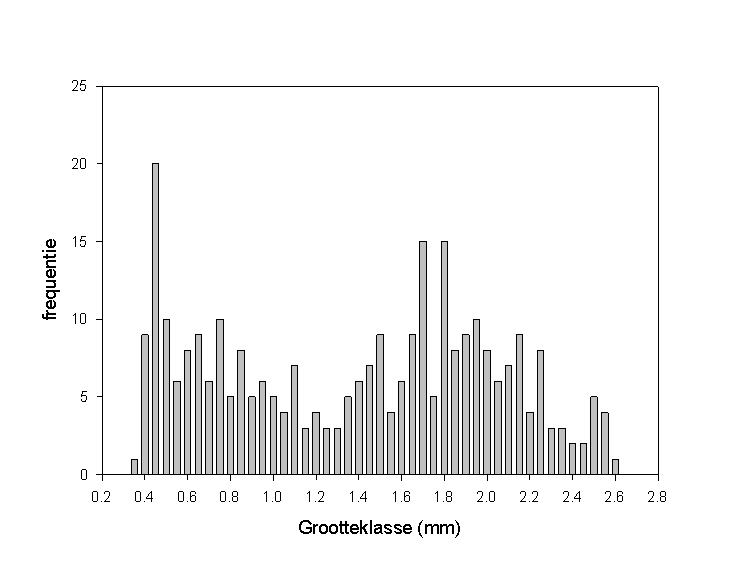
Fig. 6.6 Frequentieverdeling in functie van de lichaamslengte van Isotoma viridis.
Met uitzondering van de wintermaanden (periodes 12, 1 en 2) en periode 8 (8/7-13/8), is de vangstopbrengst groot genoeg om het gewenste aantal specimens (30 ind/periode) te meten. Daar het gebruik van de pitfall-methode een idee geeft van de locomotorische activiteit, welke door verscheidene factoren zoals vochtigheid en temperatuur beinvloed wordt (Joosse & Groen, 1970; Berbiers et al., intern rapport), heeft het aantal individuen per grootteklasse eerder een beperkte en relatieve betekenis. Niettegenstaande dit, toont de frequentieverdeling voor de opeenvolgende periodes (fig.6.7) duidelijk het voorkomen van twee gescheiden generaties.
Figuur 6.8 geeft aan dat 64% van de specimens gevangen in maart een lichaamslengte hebben tussen 1 mm en 2 mm, 13% is groter en 23% is kleiner. Deze najaarsgeneratie geeft leven aan een nieuw nageslacht vanaf mei. In deze periode kennen de adulten een snelle groei en tegen de eerste helft van mei is 43% van de gevangen specimens groter dan 2 mm, terwijl slechts 17% tot de kleinste individuen behoort. Twee maanden later is de situatie omgekeerd: er zijn slechts 17% grote individuen verzameld en 40% juvenielen (< 1 mm). De najaarscohorte is nu aan het uitsterven en wordt vervangen door de generatie uitgekomen in mei. Tegen september heeft 80% van de vangstopbrengst een lichaamsgrootte langer dan 1mm. Op dit moment is de groottesamenstelling van de vangstopbrengst nagenoeg gelijk aan deze van april.
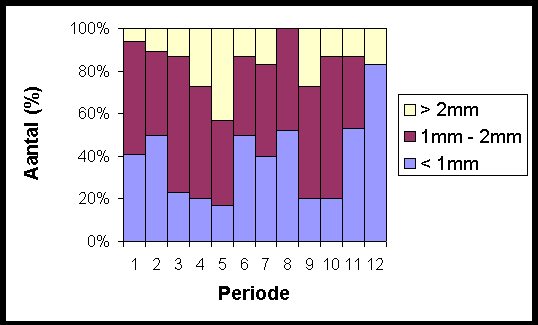
Fig. 6.8 Verdeling van de springstaarten volgens grootte over een jaar (12 periodes).
In augustus heeft geen enkel individu een lengte groter dan 2 mm. Dit kan verklaard worden doordat de najaarsgeneratie volledig uitgestorven is, terwijl de voorjaarsgeneratie nog geen grote individuen bevat. Een tweede generatie sluipt uit vanaf september en vertegenwoordigt 53% van de gevangen individuen in november.
Vanaf december is de vangstopbrengst veel lager en in december zelf konden slechts zes specimens gevangen worden, waarvan er vijf kleiner zijn dan 1 mm. Er vindt geen reproductie meer plaats en een trage groei is merkbaar gedurende januari en februari. In maart zijn opnieuw 77% springstaartjes groter dan 1 mm, terwijl deze fractie in februari slechts 50% bedraagt. De tijdelijke stop in reproductie en groei, veroorzaakt door de koude wintertemperaturen, resulteert in twee duidelijk gescheiden generaties.
Tabel 6.3 Verdeling van de springstaarten volgens grootte voor de 12 periodes, uitgedrukt in percentages.
|
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|
< 1mm |
41% |
50% |
23% |
20% |
17% |
50% |
40% |
52% |
20% |
20% |
53% |
83% |
|
1mm - 2mm |
53% |
39% |
64% |
53% |
40% |
37% |
43% |
48% |
53% |
67% |
34% |
0% |
|
> 2mm |
6% |
11% |
13% |
27% |
43% |
13% |
17% |
0% |
27% |
13% |
13% |
17% |
6.2.2 Samenvatting
Isotoma viridis, bemonsterd in de duinroosruigte, wordt gekenmerkt door twee generaties per jaar: een snel groeiende voorjaarscohorte en een traag groeiende najaarscohorte.
6.3 Verticale verspreiding
6.3.1 Bespreking
De resultaten van de niet-parametrische Mann-Whitney U-test hebben betrekking op de soorten Isotoma viridis, Entomobrya nivalis, Lepidocytus cynaeus, op de familie Poduroidea en op het totaal aantal springstaarten, verzameld op 15/5/00. Van de tien bodemstalen, verzameld op 9/9/99, zijn enkele substalen gecontroleerd op de aanwezigheid van collembolen, met een negatief resultaat als gevolg. Wanneer we bedenken dat de drie bodemstalen, genomen in de namiddag van 15/5/00 respectievelijk één en twee keer drie springstaarten bevatten, is het niet onbegrijpelijk dat de substalen geen specimens bevatten. De helft van de bodemstalen is genomen met een gutsboor die tot vijftig cm in de grond reikt en ook in deze stalen is geen leven te bespeuren. De procedure voor de staalname op 15/5/00 is dan enigszins gewijzigd, in die zin dat de staalname zowel overdag als in de late avond diende uitgevoerd te worden, om een vergelijking te kunnen maken tussen de activiteiten. Er is ook een reductie doorgevoerd wat het aantal stalen betreft.
Aangezien het beperkt aantal stalen (2�5 aspiraten en 2�3 bodemstalen), zullen de resultaten van deze steekproef geen feilloze weergave vormen van de werkelijke populatiekenmerken. Vooral de gegevens van de bodemstalen zijn niet veelzeggend. Op een, met de aspirator bemonsterd, oppervlak van (40�40) cm2 is slechts één bodemstaal genomen zodat de gevens van beide staalnametypes niet kunnen vergeleken worden. Met aspiraat worden de opgezogen specimens, met behulp van een aspirator, bedoeld. Het aantal gevangen specimens in de bodemstalen kan verdeeld worden over twee lagen: een bovenste laag tot 2.5 cm in de bodem en een onderste laag van 2.5 cm tot 6 cm diepte. De aantallen zijn zo extreem laag, dat de waarden van beide lagen gesommeerd worden (tabel B.17). De ruwe gegevens zijn beschikbaar in tabel B.18, terwijl tabel 6.4 een weergave vormt van het relatieve voorkomen van de verschillende taxa, uigedrukt in totale aantallen en in percentages.
Tabel 6.4 Vangstaantalen van de verschillende taxa per periode (dag/nacht) in het aspiraat en de bodemstalen, uitgedrukt in totalen en percentages.
|
Soort |
Aspiraat |
|
|
|
Bodemstaal |
|
|
|
|
|
Dag |
% |
Nacht |
% |
Dag |
% |
Nacht |
% |
|
Isotoma viridis |
20 |
20,6 |
257 |
67,5 |
4 |
57,1 |
9 |
19,1 |
|
Entomobrya nivalis |
41 |
42,3 |
106 |
27,7 |
1 |
14,3 |
4 |
8,5 |
|
Lepidocyrtus cynaeus |
8 |
8,2 |
0 |
0 |
1 |
14,3 |
5 |
10,7 |
|
Poduroidea |
6 |
6,2 |
1 |
0,3 |
1 |
14,3 |
20 |
42,6 |
|
rest |
22 |
22,7 |
17 |
4,5 |
0 |
0 |
9 |
19,1 |
|
totaal |
97 |
100 |
381 |
100 |
7 |
100 |
47 |
100 |
De temperatuur�en vochtigheidsmetingen zijn weergegeven in tabel 6.5. Zowel de temperatuur in de bodem als aan het bodemoppervlak ligt �s nachts lager dan overdag (figuur B.10). Tabel B.18 geeft voor beide temperatuursverschillen hoge significantiewaarden (p=0.01) weer. Ook de luchtvochtigheid vertoont hoog significante verschillen, terwijl dit niet het geval is voor de bodemvochtigheid. De gemiddelde luchtvochtigheid �s nachts is meer dan het dubbele van de gemiddelde waarde overdag.
Tabel 6.5 Temperatuur- en vochtigheidsmetingen .
|
|
bodemtemperatuur (�C) |
bodemvochtigheid |
temperatuur (�C) |
luchtvochtigheid |
||||
|
|
dag |
nacht |
dag |
nacht |
dag |
nacht |
dag |
nacht |
|
aantal |
29,9 |
19,9 |
77,6 |
95,5 |
33,8 |
17 |
57,6 |
77,5 |
|
|
32,2 |
17,2 |
91,3 |
99,9 |
29,3 |
14,3 |
45,2 |
85,4 |
|
|
32,6 |
17,9 |
97,3 |
99,9 |
34,1 |
14,7 |
36,6 |
97,7 |
|
|
32 |
17,9 |
95,3 |
99,9 |
35,9 |
14,4 |
35,4 |
97,4 |
|
|
30 |
20,5 |
99,9 |
99,6 |
38,4 |
14,7 |
38,3 |
94,3 |
|
gem. |
31,34 |
18,68 |
92,28 |
98,96 |
34,3 |
15,02 |
42,62 |
90,46 |
Een eerste blik op tabel 6.6 laat ons zien dat het merendeel van de onderzochte dag-nacht verschillen veroorzaakt wordt door fluctuaties als gevolg van een random staalname (NS). De bekomen resultaten geven enkel een significant dag-nacht-verschil (p=0.049) weer voor het totaal aantal collembolen verzameld met de bodemstalen. Uit figuur B.11 blijkt dat het aantal gevangen specimens �s avonds veel hoger ligt dan overdag. Wanneer we de soorten afzonderlijk beschouwen zijn geen significante verschillen merkbaar (figuren: zie bijlage: B.12-B.21). De juist op de grens liggende significantiewaarde voor het totaal aantal collembolen kan begrepen worden door het feit dat voor alle betrokken soorten, de vangst met behulp van bodemstalen �s avonds steeds groter (maar niet significant) is dan overdag. Anderzijds is ook het beperkt aantal stalen een invloedrijke factor. In vergelijking met de andere soorten, zijn het voornamelijk de Poduroidea die �s avonds veel meer in de bodem aanwezig zijn dan overdag (tabel 6.4). Zij zullen dan ook de grootste invloed uitoefenen op de totale verschillen.
Wanneer we de aspiraten beschouwen, is er geen enkel verschil dat significant is. Nochtans deden de hoge aantallen van Isotama viridis tijdens de nachtelijke uren een beduidend verschil tussen de gemiddelden vermoeden. Het probleem kan snel begrepen worden wanneer we de ruwe data er bij nemen en zien dat één van de vijf waarnemingen uitzonderlijk laag is in vergelijking met de andere (tabel B.18). We kunnen deze waarde beschouwen als een uitbijter of misschien is er iets misgelopen tijdens de staalname zelf, want ook het totaal aantal gevangen Collembola is opvallend laag. Na het verwijderen van de desbetreffende waarde wordt een significante p-waarde (p=0.014) verkregen voor het verschil tussen de gemiddelde aantallen van Isotoma viridis in het aspiraat (tabel 6.6*). Nochtans is op figuur B.13 nog steeds een overlapping merkbaar, welke het gevolg is van de �s avonds voorkomende grote spreiding van de waarden. Deze figuur toont ook dat het relatieve voorkomen van Isotoma viridis aan de oppervlakte �s avonds veel groter is. Ook in tabel 6.4 is te zien dat de vangst �s avonds beduidend groter is dan overdag.
Tabel 6.6 Weergave van het significantieniveau voor de verschillen in het relatief voorkomen van de springstaarten overdag en �s nachts. NS*: na het verwijderen van de uitbijter is de p-waarde significant (p=0.014, S). De significante p-waarde voor het totaal aantal springstaarten bedraagt 0.049.
|
|
Aspiraat |
|
Bodemstaal |
|
|
|
Dag |
Nacht |
Dag |
Nacht |
|
Isotoma viridis |
NS* |
NS* |
NS |
NS |
|
Entomobrya nivalis |
NS |
NS |
NS |
NS |
|
Lepidocyrtus cynaeus |
NS |
NS |
NS |
NS |
|
Poduroidea |
NS |
NS |
NS |
NS |
|
Totaal |
NS |
NS |
S |
S |
Net zoals bij de bodemstalen, is ook hier het aantal gevangen collembolen �s avonds hoger dan overdag. De Poduroidea en Lepidocyrtus cynaeus vormen de enige uitzonderingen op het globale patroon: de aspiraten van de namiddag bevatten meer specimens dan deze van de avond. Maar deze niet-significante verschillen hebben weinig of geen betekenis.
6.3.2 Samenvatting
Algemeen kan gesteld worden dat collembolen in de mosduinen �s nachts veel talrijker zijn dan overdag. Van het totaal aantal gevangen springstaarten tijdens de nacht, is slechts 24% terug gevonden overdag. De verwachte hypothese dat collembolen zich gedurende de warme namiddag voornamelijk in de bodem schuilhouden en �s avonds, wanneer de luchtvochtigheid toeneemt en de temperatuur afneemt, opnieuw naar de oppervlakte begeven, blijkt niet op te gaan voor deze studie. Integendeel, zo is voor het totaal aantal collembolen aangetoond dat ze tijdens de koelere en vochtigere avonduurtjes meer in de diepere bodemlagen voorkomen dan overdag. Isotoma viridis en Entomobrya nivalis vertonen een gelijkaardig patroon; �s nachts bevinden ze zich bijna hoofdzakelijk aan het bodemoppervlak, terwijl in de namiddag geen beduidende verschillen kunnen waargenomen worden, daar het aantal gevangen specimens dan respectievelijk slechts 9 en 38% van de nachtelijke vangst bedraagt. Het tegengestelde geldt voor de twee overige behandelde taxa, de Poduroidea en Lepidocyrtus cynaeus, die �s nachts meer de voorkeur bieden aan de diepere, minerale bodemlagen.
6.4 Horizontale verspreiding en predator- prooi correlatie
6.4.1 Bespreking
Tabel B.20 geeft het aantal gevangen collembolen en spinnen weer. De minder talrijke soorten, waaronder Orchesella villosa, Cryptopygus thermophilus, Xenylla maritima, Neanura muscorum en Parisotoma notabilis, worden als restgroep beschouwd.
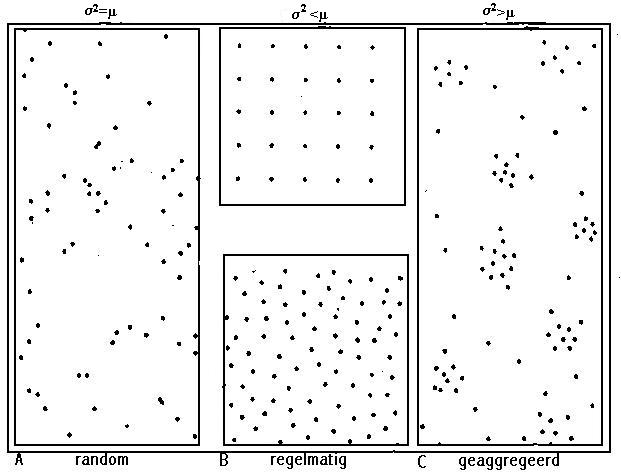
Fig. 6.9 Agregatietypes.
Bij een gegeven gemiddelde densiteit (x), wordt de mate van aggregatie van de te onderzoeken soort weergegeven door de variantie (s�) van de vangstaantallen tussen de verschillende staaleenheden (31). Men spreekt van een geaggregeerde verspreiding wanneer het gemiddelde kleiner is dan de variantie. Wanneer deze twee grootheden gelijk zijn, is er sprake van een toevallige dispersie en bij een gemiddelde groter dan de variantie, wordt een regelmatige verspreiding bekomen (fig.6.9). De verwachte aggregaatvorming kan aangetoond worden bij de drie meest voorkomende soorten, namelijk Isotoma viridis, Entomobrya nivalis en Orchesella cincta.
Tabel B.20 en B.21 tonen dat Isotoma viridis en Entomobrya nivalis zowel in de habitatten afzonderlijk (A, B en C) als samen (totaal) aggregaten vormen, terwijl Orchesella cincta in habitat B en C geen aggregatie vertoont (tabel B.22), maar een uniforme tot zelfs toevallige verspreiding. Wanneer de verhouding x/s2 bepaald wordt, merken we dat alle drie de soorten de grootste aggregatie vertonen in habitat A (duingrasland). Isotoma viridis is het minst geaggregeerd in habitat B (kruidenvegetatie), terwijl bij Entomobrya nivalis de aggregatie het minst uitgesproken is in habitat C (mosduin), waar de verspreiding van Orchesella cincta zelfs toevallig is. Een volledige beschrijving van de habitatten is terug te vinden in punt 5.1. Hier zullen we de meetresultaten van de verschillende bodemlagen overnemen (tabel 6.7). De gegevens van habitat D en E zijn niet bruikbaar voor het bepalen van aggregaten, daar het determineren en tellen niet per staal is gebeurd maar per habitat.
Tabel 6.7 Meetresultaten van de fragmentatielaag en bodemontwikkeling en weergave van de aan- of afwezigheid van een strooisellaag voor de vijf microhabitatten.
|
|
A |
B |
C |
D |
E |
|
strooisel |
- |
+ |
+ |
- |
+ |
|
F-laag |
1 cm |
1.5 - 2 cm |
3 mm |
3 mm |
1 cm |
|
B-O |
20 - 25 cm |
20 - 25 cm |
2 - 3 cm |
2 - 3 cm |
3 - 3.5 cm |
Vervolgens kunnen we stellen dat het relatieve voorkomen van Isotoma viridis en Orchesella cincta vrij gelijklopend is voor de drie habitatten, terwijl Entomobrya nivalis beduidend meer voorkomt in habitat A dan C (tabel B.23, B.24 en fig.B.22). De grafieken van de niet-significante variantie tussen de habitatten zijn terug te vinden in de bijlagen, fig.B.23 en B.24. Daar van habitat D en E enkel gegevens over het aantal individuen per habitat beschikbaar zijn, kunnen deze data wel gebruikt worden om veronderstellingen te maken maar niet om variantieanalyse uit te voeren. In tabel 6.8, die het gemiddeld aantal individuen per staal weergeeft, is te zien dat het voorkomen van Entomobrya nivalis in habitat D nog lager is dan in habitat C. Wanneer we weten dat het verschil in relatief voorkomen tussen habitat A en C significant is, kunnen we met grote waarschijnlijkheid veronderstellen dat dit tussen habitat A en D ook significant zal zijn. De hoge aantallen van Entomobrya nivalis in habitat A zijn niet uitzonderlijk; figuur 6.10 laat zien dat ook het totaal aantal gevangen collembolen het hoogst is in A.
Tabel 6.8 Het gemiddeld aantal individuen per staal voor de vijf microhabitatten.
|
|
A |
B |
C |
D |
E |
|
Isotoma viridis |
35,2 |
27,1 |
29,1 |
21,4 |
26,8 |
|
Entomobrya nivalis |
15,8 |
12,2 |
6,2 |
3,6 |
7,6 |
|
Orchesella cincta |
1,1 |
1 |
1,3 |
0 |
0 |
|
rest |
1,1 |
0,8 |
3,2 |
0,4 |
0 |
|
totaal |
53,2 |
41,1 |
39,8 |
25,4 |
34,4 |
De oorzaak voor het relatief lage voorkomen in habitat C kan waarschijnlijk gezocht worden in de aanwezigheid van een uiterst dunne fragmentatielaag (3 mm) en de weinige bodemontwikkeling (2 à 3 cm), waardoor er minder vocht wordt vastgehouden. De invloed van de strooisellaag wordt nagegaan door een vergelijking te maken tussen habitat C en D. In tabel B.25 kunnen we zien dat onze hypothese correct is: een hoog significante correlatie wordt bekomen voor het relatieve voorkomen van Entomobrya nivalis en de mate van bodemontwikkeling (r=0.98, p=0.005)(fig.B.25). De correlatie met de dikte van de F-laag is vrij hoog maar niet significant, zodat ook de correlatie met de twee lagen samen hoog is maar net niet significant (fig.B.26). Voor de correlatie tussen de bodemontwikkeling en het totaal aantal specimens wordt eveneens een hoge waarde verkregen (r= 0.82), maar zoals op figuur B.27 te zien is, is deze niet significant. Verder is ook het verband met het percentage mosbedekking nagegaan, maar geen van de onderzochte gevallen is significant. Wel is in drie van de vier gevallen het aantal individuen omgekeerd gecorreleerd met de percentuele mosbedekking. Het algemeen lagere voorkomen in habitat D, wijst erop dat de afwezigheid van de strooisellaag haar invloed uitoefent. Toch is deze afwezigheid hier niet de oorzaak van het relatief lagere voorkomen van de collembolen, want ook habitat A bezit geen strooisellaag. Het is eerder een combinatie van de afwezigheid van een strooisellaag en een beperkte bodemontwikkeling in een open mosduinvlakte die verantwoordelijk is voor het relatief lage voorkomen.
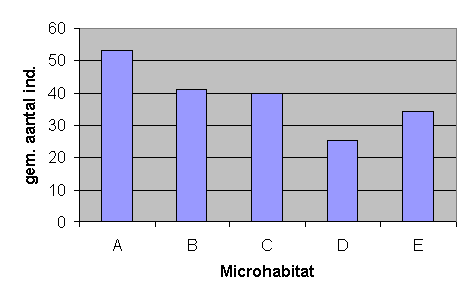
Fig. 6.10 Het gemiddeld aantal springstaarten per microhabitat.
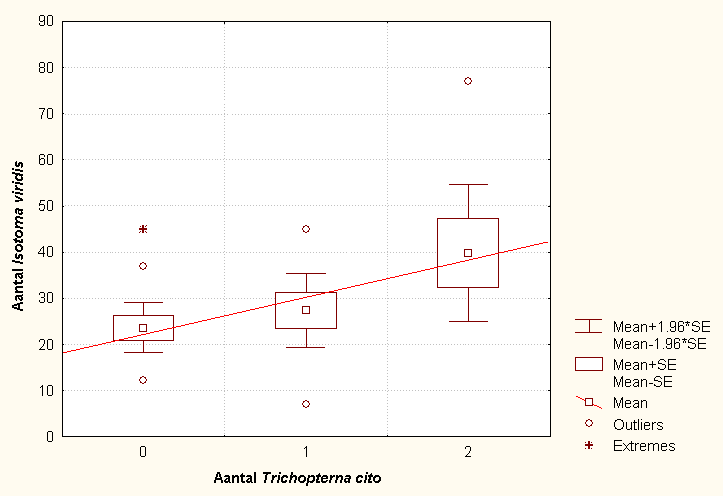
Naast de collembolen werden ook de spinnen uit de pitfalls gedetermineerd en geteld. Tabel B.26 geeft een overzicht van de resultaten. Opnieuw zijn de gegevens van habitat D en E niet bruikbaar, zodat er in totaal 31 steekproeven zijn (N). De spinnen behoren tot 18 verschillende soorten, waarvan Trichopterna cito (Linyphiidae) en Pardoa monticola (Lycosidae) als belangrijke predators van de Collembola kunnen beschouwd worden. Een van de onderzochte variabelen is het totaal aantal spinnen verminderd met het aantal gevangen specimens van Alopecosa barbipes (Lycosidae), omdat het hier om een vrij grote soort gaat. In tabel B.28 en figuur 6.11 zien we dat er enkel een lage, significante correlatie (p=0.021, r=0.417) bestaat tussen het relatieve voorkomen van Trichopterna cito en Isotoma viridis.
Fig. 6.11 Grafiek die de correlatie tussen Trichopterna cito en Isotoma viridis weergeeft.
6.4.2 Samenvatting
We kunnen besluiten dat de onderzochte mate van aggregatie het kleinst is bij Orchesella cincta. Vervolgens hebben de verschillende bodemkenmerken weinig invloed op de horizontale verspreiding van Isotoma viridis, terwijl Entomobrya nivalis opvallend minder voorkomt in de mosduinvlakte waar de bodemontwikkeling minimaal is. Tenslotte stellen we vast dat er een verband bestaat tussen het relatieve voorkomen van Trichopterna cito en Isotoma viridis.
Go to the
first,
previous,
next,
last
chapter,
overview.